7
Mentes sexuadas
Como hemos visto en los capítulos precedentes, la selección natural primero y la selección sexual después tienden a exacerbar la asimetría macho-hembra y, por ende, el dimorfismo sexual (sea éste anatómico, fisiológico o conductual). Aunque a menudo la selección sexual se traduce en un mayor tamaño del sexo masculino, como ocurre en la mayoría de mamíferos y aves sexualmente dimórficos, en realidad esto es más la excepción que la regla. En la mayoría de especies de insectos, arácnidos, peces, anfibios y serpientes, las hembras son mayores que los machos. (Incluso entre los mamíferos, los casos de mayor tamaño femenino son menos raros de lo que podría pensarse: las hienas, los conejos y liebres, una familia de murciélagos, tres familias de cetáceos, un grupo de focas y dos tribus de antílopes cuentan con especies cuyas hembras son mayores que los machos). No es que ser más grande que los rivales ya no sea una ventaja para los machos de estas especies, lo que ocurre es que también las hembras más grandes dejan más descendencia que las otras, sea porque producen más crías o porque éstas nacen más rollizas. Además, la producción de huevos suele requerir más espacio y recursos (y, por ende, un cuerpo más grande) que la producción de espermatozoides diminutos y sin sustancias de reserva. Esto vale sobre todo para los invertebrados, pero incluso entre los vertebrados encontramos casos extremos de este dimorfismo de tamaño invertido (desde nuestra perspectiva mamiferocéntrica), como el de los peces del género Ceratias. Estos pejesapos abisales, que alcanzan el metro de longitud, flotan inmóviles en la oscuridad de las aguas profundas mientras balancean una espina modificada luminiscente a modo de cebo para atraer presas. Los primeros ictiólogos que los describieron, allá por los años veinte del siglo pasado, repararon en que casi todos los ejemplares que capturaban, siempre hembras, llevaban adheridos a la piel lo que parecían ser alevines parásitos. Pero pronto comprobaron que no se trataba de especímenes inmaduros, sino de machos adultos diminutos y «degenerados» que vivían permanentemente fusionados a la hembra, de la que se habían convertido en un apéndice, formando ambos sexos una suerte de superorganismo hermafrodita.
Si una desigualdad sexual tan exagerada puede darse en la naturaleza es precisamente porque ambos sexos son iguales a ojos de la selección natural. Como he remarcado en el capítulo 3, en su tránsito de una generación a la siguiente los genes pueden acceder indistintamente a un vehículo masculino o femenino (salvo una pequeña minoría de genes que se perpetúan sólo por vía materna o paterna, cuyo interés egoísta en favorecer un sexo a expensas del otro debe supeditarse al interés de la mayoría neutral). Por eso las relaciones de dominancia o dependencia entre los sexos derivadas del dimorfismo sexual son irrelevantes para la selección natural.
Aun así, el dimorfismo sexual extremado es bastante raro. Los machos y hembras de la mayoría de especies animales son muy similares, si no indistinguibles. Como es natural, las diferencias más obvias pertenecen al dominio reproductivo, donde la masculinidad y la feminidad están claramente delimitadas[129]. Pero incluso en este terreno hay cierta discrepancia. Por supuesto, las diferencias en la anatomía y la fisiología reproductivas suscitan poca controversia, pero no ocurre lo mismo con las diferencias de comportamiento, cuya determinación genética es mucho más discutible. Algunas feministas han llegado a cuestionar la existencia de una inclinación maternal femenina innata, y esgrimen los resultados de algunos experimentos clásicos con macacos privados de contacto social. Las monas criadas sin ninguna interacción social con sus congéneres no sólo tenían dificultades para aparearse (desconocían los códigos, las respuestas y las posturas sexuales propias de su especie) sino que, si se las fecundaba por inseminación artificial, las madres primerizas hacían caso omiso de la cría recién nacida o, peor aún, la veían como un animal extraño, llegando incluso a atacarla (aunque hay que decir que su competencia como madres iba mejorando con la experiencia, de manera que los hijos posteriores tenían más posibilidades de salir adelante)[130]. Pero entre machos y hembras hay una diferencia significativa, y es que, cuando las monas juveniles crecen en un entorno social normal, muestran gran curiosidad por los bebés y los cuidados que les dispensan sus madres, que intentan imitar siempre que tienen ocasión, un interés que no exhiben (o lo hacen en mucha menor medida) los machos de la misma edad. La adquisición del comportamiento maternal de las hembras primates parece requerir un entorno social propicio durante un periodo crítico, pero lo mismo ocurre con el lenguaje humano, y no por ello pondríamos en duda que la diferencia de capacidad lingüística entre chimpancés y seres humanos tiene una base genética. La expresión de los genes siempre es contextual, de modo que la ausencia de un rasgo (conductual o de otra naturaleza) en contextos artificiales o anómalos no excluye su determinación genética (aunque sea sólo parcial).
Cerebros masculinos y cerebros femeninos
Los machos reconocen a las hembras fecundables, se exhiben e insinúan ante ellas y (si tienen suerte) las inseminan, mientras que las hembras seleccionan aspirantes a padre, solicitan la cópula y cuidan de las crías. Estas diferencias conductuales tan generalizadas (aunque ni mucho menos universales) entre los sexos han llevado a muchos a pensar que machos y hembras deben diferir esencialmente en aspectos clave de la circuitería y/o la función de la maquinaria responsable del comportamiento: el sistema nervioso central.
Desde los años setenta del siglo pasado han proliferado los informes de dimorfismos sexuales en el cerebro tanto de vertebrados como de invertebrados, que incluyen diferencias en el tamaño cerebral total, el tamaño de regiones o subregiones cerebrales concretas, el número de neuronas o su tamaño en ciertas regiones cerebrales, la ramificación dendrítica de las neuronas, la organización sináptica, la estructura de la membrana neuronal e incluso la distribución y concentración relativa de receptores de neurotransmisores, receptores de hormonas esteroides, actividades enzimáticas o proteínas citoesqueléticas[131]. Hoy se asume que los dimorfimos sexuales cerebrales son ubicuos en el reino animal, aunque no hay tanto acuerdo en lo que respecta a los mecanismos de la diferenciación sexual del cerebro y la traducción de las diferencias cerebrales en diferencias comportamentales.
Además de los genes, el entorno y el comportamiento influyen en la diferenciación sexual del cerebro. Por ejemplo, en un experimento se colocaron ratas recién destetadas en entornos monótonos o entretenidos y se examinó el desarrollo cerebral de los machos y las hembras al cabo de un mes. Las ratas que habían crecido en jaulas de laboratorio ordinarias (un entorno bastante aburrido) apenas exhibían diferencias de comportamiento. En cambio, los machos que habían crecido en entornos más variados, dotados de aparatos para la estimulación sensorial y el juego, exhibían comportamientos ausentes en las hembras, así como un córtex visual con mayor ramificación dendrítica en comparación tanto con las hembras criadas en las mismas condiciones como con los machos criados en el entorno sin estímulos. (La interpretación evolutiva de este dimorfismo sexual dependiente del entorno es oscura. No se sabe por qué los cerebros de las ratas de ambos sexos responden de manera distinta a los estímulos externos, ni se conocen los factores mediadores de la diferenciación cerebral inducida por el entorno)[132].
El cerebro embrionario, inicialmente asexual, se desarrolla en el sentido femenino o masculino según la pauta de secreción de hormonas ováricas o testiculares, su conversión local y la localización y cantidad de receptores celulares de hormonas esteroides. La expresión de dichos receptores y conversiones enzimáticas en ciertas regiones cerebrales influye en parámetros tales como el número de neuronas. Así, un cambio en la densidad de neuronas de una o varias regiones puede traducirse en diferencias de conectividad o densidad sináptica, propiedades electrofisiológicas o supervivencia neuronal en dichas regiones y sus conexiones aferentes y eferentes. La acción subsiguiente de nuevas oleadas de hormonas esteroides refuerza este dimorfismo sexual incipiente. Los estadios iniciales de la diferenciación cerebral parecen ser impermeables a las influencias externas, pero el desarrollo posterior sí depende de factores físicos, ambientales y sociales (edad, rango, estrés, nutrición) que alteran la producción de hormonas sexuales y pueden así determinar el destino ulterior de los circuitos cerebrales sexualmente dimórficos.
Dimorfismo cerebral y conducta
Las diferencias de comportamiento entre machos y hembras son de dos tipos. Hay conductas que son exclusivas de un sexo y están ausentes en el otro. Ejemplos triviales son la penetración masculina y el parto o la puesta de huevos. Muchas exhibiciones de cortejo y cantos para atraer parejas sexuales son exclusivos de un sexo (casi siempre el masculino). También buena parte del comportamiento de crianza puede ser exclusivo de un sexo (casi siempre el femenino). Pero las conductas exclusivamente masculinas o femeninas suelen representar sólo una pequeña fracción del repertorio conductual de ambos sexos. Casi todas las conductas (incluidas las sexuales) pueden darse en los dos sexos, aunque con más frecuencia en uno que en otro, en cuyo caso se habla de comportamiento típicamente masculino o femenino. Esto puede deberse a que machos y hembras difieran en sus predisposiciones, o a que, aunque sean igualmente proclives a la conducta en cuestión, las situaciones particulares que la propician son más frecuentes para un sexo que para el otro. (Un ejemplo es la caza en los chimpancés, practicada mayoritariamente por bandas de machos, con la ocasional incorporación de hembras estériles).
Los comportamientos típicos de un sexo son problemáticos, porque el dimorfismo conductual puede depender de la situación. Por ejemplo, un macaco macho que se encuentre solo responderá a las llamadas de angustia de una cría aislada tal como lo haría una hembra. Pero si hay alguna hembra presente, permanecerá impasible y dejará que sea ella quien se ocupe de la cría. En este caso, como en muchos otros, el dimorfismo conductual sólo se manifiesta en ciertos contextos sociales. En el pinzón cebra, la conducta típicamente femenina de solicitud de la cópula consiste en agacharse y arquear el cuerpo para presentar la cloaca al macho, junto con un aleteo rápido y vocalizaciones características. Pero esta misma conducta también se observa en los machos tras la cópula, y se especula que en este caso podría tener una función social bien diferente. El amplio solapamiento entre los repertorios conductuales de machos y hembras también dificulta la tarea de relacionar dimorfismos cerebrales con diferencias de conducta. Por otra parte, en algunas especies no todos los individuos optan por la misma estrategia de apareamiento. Algunos primates, como el papión gelada, combinan la poliginia de harén con la monogamia. En otras especies hay dos tipos de machos, unos «viriles» y otros de aspecto y modales femeninos (como ocurre, por ejemplo, en algunos peces cuyos machos «afeminados» se inmiscuyen en el apareamiento de un rival «viril» para verter su semen subrepticiamente y fecundar parte de los huevos recién depositados por la hembra).
Aun así, numerosos estudios han permitido correlacionar diferencias de comportamiento entre machos y hembras con estructuras cerebrales o pautas endocrinas sexualmente dimórficas. Por ejemplo, el núcleo dimórfico de la región preóptica del hipotálamo se correlaciona con el comportamiento copulatorio masculino y femenino de la rata, y el dimorfismo en la arquitectura endocrina del hipotálamo ventromedial se correlaciona con la ausencia de lordosis en la rata macho. Igualmente, el dimorfismo de la región preóptica medial se correlaciona con diferencias sexuales en el comportamiento odorífero de los jerbos, y el dimorfismo de tamaño y densidad neuronal en los núcleos frontales del canto se correlaciona con diferencias sexuales en las vocalizaciones de los canarios y otras aves canoras. Pero, más allá de estas diferencias generales, es muy difícil atribuir consecuencias conductuales precisas a diferencias cerebrales entre machos y hembras individuales.
A menudo, ni siquiera las diferencias de comportamiento sexual pueden correlacionarse con diferencias cerebrales apreciables. Un ejemplo significativo lo encontramos en unos reptiles que se han pasado a la reproducción asexual, pero que continúan exhibiendo comportamientos copulatorios: las lagartijas partenogenéticas del género Cnemidophorus (las madres vírgenes lesbianas que han inspirado el título de este libro). Como hemos visto en el capítulo inicial, se trata de una especie unisexual formada exclusivamente por hembras (en el sentido de que poseen ovarios en vez de testículos, y ponen huevos). Y aunque no necesitan del sexo para reproducirse, continúan practicándolo en una versión «lésbica» de la cópula heterosexual. Lo que nos interesa aquí es que, mientras que algunas hembras adoptan el rol «activo» en estos simulacros de cópula, otras se dejan «montar», dependiendo de la fase del ciclo ovulatorio (las hembras adoptan una conducta sexual típicamente femenina cuando se avecina la ovulación, mientras que el resto del tiempo se comportan como lo haría un macho). Pues bien, aunque las especies dioicas emparentadas presentan un dimorfismo sexual reseñable en ciertas regiones cerebrales implicadas en la conducta sexual, las lagartijas unisexuales que se comportan como machos o como hembras no exhiben ningún dimorfismo cerebral comparable[133].
Por otra parte, el dimorfismo cerebral no siempre se traduce en diferencias manifiestas de conducta o aptitudes entre machos y hembras. Los neurólogos suelen dar por sentado que la extensión de una región cerebral se correlaciona con la complejidad del comportamiento controlado por la misma. En el caso de los pájaros canoros, por ejemplo, se supone que el mayor desarrollo de las áreas del canto en los machos tiene que ver con la mayor complejidad o variedad de su repertorio sonoro. Pero esto sólo vale para las especies cuyas hembras cantan poco o nada. Cuando ambos sexos son igual de cantarines, a menudo el dimorfismo cerebral no parece entrañar ninguna diferencia obvia de aptitud canora. Por ejemplo, en las cosifas africanas, que ejecutan duetos, las áreas del canto masculinas son mayores que su contrapartida femenina, a pesar de lo cual la hembra es perfectamente capaz de interpretar el dueto entero cuando se la aísla de su pareja. En el caso del canario, las hembras a las que se administra testosterona exhiben la misma aptitud canora que los machos, aunque sus áreas de canto sólo contienen la mitad de neuronas. Hasta las hembras de pinzón cebra, que en condiciones normales no cantan en absoluto, sometidas al mismo tratamiento hormonal demuestran estar casi tan dotadas para el canto como los machos, a pesar de que éstos tienen áreas de canto entre tres y cuatro veces más extensas[134].
Incluso los dimorfismos que afectan a regiones cerebrales directamente implicadas en la conducta sexual pueden resultar difíciles de interpretar. En la rata hembra, el núcleo sexualmente dimórfico del área preóptica medial está atrofiado, lo que se ha ligado a la ausencia de comportamiento copulatorio «activo». Pero las lesiones de esta región cerebral no parecen perturbar en absoluto la conducta copulatoria de los machos sexualmente experimentados. En el caso de los machos vírgenes, la única manifestación es una falta de respuesta sexual ante hembras poco tentadoras. Las lesiones del núcleo sexualmente dimórfico no afectan las respuestas hormonales masculina o femenina a la estimulación sexual, ni siquiera en los individuos vírgenes. Así pues, no parece que esta región cerebral sexualmente dimórfica sea indispensable para la plena expresión del comportamiento copulatorio (aunque sí parece ejercer cierta influencia en la excitación sexual masculina). No está claro, por lo tanto, que el tamaño del núcleo sexualmente dimórfico tenga algo que ver con la variabilidad individual de la conducta sexual[135].
El dichoso cuerpo calloso
Los dimorfismos cerebrales documentados en ratas, pájaros y otros animales desataron en su momento una auténtica fiebre del oro entre los neurólogos ávidos de encontrar un correlato cerebral de las presuntas diferencias psicológicas entre varones y mujeres. Aunque la afirmación de los craneómetras decimonónicos de que los varones tenían cerebros mayores que las mujeres (y, por ende, eran más inteligentes) hace tiempo que ha quedado desacreditada (véase la figura 7.1), los ahora más sofisticados buscadores de diferencias cerebrales entre ambos sexos no han dejado de encontrarlas. La larga lista incluye estructuras mayores en el cerebro masculino (el núcleo de la estría terminal, los núcleos intersticiales segundo y tercero del hipotálamo anterior, el núcleo sexualmente dimórfico del área preóptica), estructuras mayores en el cerebro femenino (la comisura anterior, el cuerpo calloso, el istmo del cuerpo calloso, la masa intermedia del tálamo) y diferencias de forma (un esplenio del cuerpo calloso más bulboso en las mujeres, y un núcleo supraquiasmático más esférico en los varones)[136].
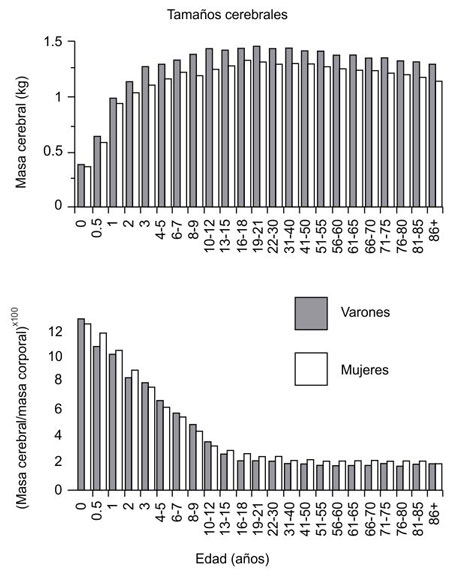
Figura 7.1. Tamaños cerebrales absolutos y relativos de varones y mujeres. Como se aprecia en la gráfica, la diferencia aparente de volumen cerebral entre ambos sexos desaparece cuando se considera el tamaño cerebral en relación al tamaño corporal. En todo caso, son las mujeres las que tienen cerebros proporcionalmente algo mayores que los varones, aunque la diferencia no es significativa y puede explicarse por simple alometría. (Dekaban y Sadowsky, 1978).
También se ha dicho que el cerebro masculino es más asimétrico que el femenino o, en otras palabras, que está más lateralizado, con mayor especialización de los hemisferios. La asimetría de los hemisferios cerebrales tiene que ver con la cuestión del tamaño del cuerpo calloso (el haz de fibras nerviosas que conecta los hemisferios izquierdo y derecho), seguramente el tema que ha suscitado más controversia y más ha llamado la atención de la divulgación científica sensacionalista, hasta tal punto que la afirmación de que el cuerpo calloso es más voluminoso en la mujer que en el varón, lo que implicaría que los hemisferios cerebrales de las mujeres estarían más interconectados (un argumento que se ha esgrimido para justificar supuestas diferencias entre las psicologías masculina y femenina), casi se ha convertido en un «hecho» para el gran público. Pero lo cierto es que el entusiasmo inicial ha dado paso al escepticismo y, tras hacer balance, los expertos en el tema han acabado por admitir que no hay evidencias anatómicas objetivas de diferencias significativas entre varones y mujeres en lo que respecta al tamaño o la forma del cuerpo calloso[137].
¿Las matemáticas son cosa de hombres?
Una de las diferencias psicológicas entre varones y mujeres que algunos han querido derivar de un ilusorio dimorfismo cerebral tiene que ver con la aptitud matemática. Éste es uno de los campos de batalla principales en las discusiones sobre presuntas diferencias intelectuales innatas entre los sexos. No en vano el talento matemático suele contemplarse como la más noble de las facultades intelectuales, a menudo imbuida de un aura casi mística que va más allá de las capacidades reales en juego. Los duchos en matemáticas suelen ser considerados más inteligentes que el resto de la humanidad, aunque carezcan de talento artístico, musical o literario. Y como otras cualidades «superiores», la aptitud matemática se presupone más propiamente masculina que femenina.
El caso es que los tests psicológicos parecen abonar este extendido prejuicio sexista. Los resultados de numerosos estudios han evidenciado que los niños comienzan a aventajar a las niñas en aptitud matemática ya desde una edad bastante temprana. Pero esto no significa que la diferencia sea innata. Hay que decir que la diferencia entre las puntuaciones medias de ambos sexos no sólo depende del test aplicado, sino que ha disminuido en las últimas décadas, lo que sugiere que hay otros factores en juego aparte de las aptitudes innatas. Por ejemplo, las encuestas a los progenitores revelan que tanto las madres como los padres esperan más de sus hijos varones que de sus hijas en lo que a aptitud matemática se refiere, con independencia del rendimiento escolar de unos y otras[138].
Aunque la diferencia entre las poblaciones masculina y femenina es siempre pequeña y la variación individual amplia, por lo que no es nada raro encontrar mujeres más competentes en matemáticas que la gran mayoría de varones, algunos pedagogos se han apresurado a sugerir que ciertos temas quizá requieran una enseñanza adaptada a cada sexo para que alumnos y alumnas los comprendan igual de bien. Asimismo se ha insinuado que las jóvenes aprenden mejor en una atmósfera más cooperativa y menos competitiva. Incluso se han concebido juegos de ordenador pretendidamente destinados al público femenino, donde las tramas de relaciones sustituyen la caza de marcianos. Todas estas propuestas, seguramente bienintencionadas, asumen de manera implícita (y acrítica) la existencia de diferencias psicológicas intrínsecas entre los sexos. Pero la mayoría de expertos elude expresar esta convicción abiertamente, quizá porque, a diferencia del estudio de los dimorfismos sexuales psicológicos y neurológicos en animales no humanos, el debate científico sobre las posibles diferencias intelectuales entre varones y mujeres se enmarca en un acalorado debate ideológico, y plantear siquiera la posibilidad de que tales diferencias sean «naturales» en mayor o menor medida es políticamente incorrecto.
El problema es que ningún test psicológico puede zanjar la cuestión de hasta qué punto las diferencias individuales en la aptitud matemática (o musical, o verbal, o lo que sea) son innatas o adquiridas. En el caso que nos ocupa, podría decirse que una diferencia psicológica entre varones y mujeres tiene una base genética si la única diferencia entre ambos grupos fuera su genotipo XX o XY, con cualquier influencia del entorno invariante. Pero esta condición es impracticable. Por eso es imposible separar la naturaleza de la crianza. No obstante, aunque no es factible educar una población de niños y niñas en entornos idénticos, los estudios interculturales indican que la crianza tiene una influencia capital en el desarrollo de las aptitudes innatas. Por ejemplo, se sabe que los niños japoneses también superan a las niñas en los tests de aptitud matemática. Pero en éste y otros casos la diferencia intersexual resulta ser menor que la diferencia intercultural, de manera que las niñas japonesas aventajan a los niños norteamericanos de la misma edad por un margen más amplio que la diferencia entre éstos y sus compañeras de clase[139].
Mujeres desorientadas
Una componente principal de la pretendida diferencia de aptitud matemática entre varones y mujeres es el razonamiento espacial, o la visualización de problemas geométricos en dos o tres dimensiones. De hecho, los problemas con los que las niñas suelen tener más dificultades que los niños tienen que ver con la rotación mental de un objeto tridimensional dibujado. Esta diferencia se ha hecho extensiva a veces a la lectura de mapas o a la capacidad de orientación. Numerosos estudios han intentado evaluar la magnitud de este dimorfismo sexual psicológico. Hay que decir que también aquí la diferencia entre varones y mujeres depende del test aplicado, y en ocasiones de la edad de los sujetos. Como ocurre con la aptitud matemática general, en el caso particular de la aptitud espacial también se aprecia una reducción de dicha diferencia en los últimos años[140].
Como ya he advertido, los resultados de un test psicológico no demuestran en absoluto que la diferencia mental entre varones y mujeres sea innata, lo que no ha impedido que bastantes antropólogos y psicólogos evolucionistas se hayan lanzado a especular sobre el significado evolutivo de la mayor aptitud espacial masculina y a proponer explicaciones adaptacionistas que invocan nuestro pasado de cazadores-recolectores. Para estos autores, si dicha diferencia es una herencia de nuestros ancestros prehistóricos y tiene un significado adaptativo plausible, entonces es tan natural e inexorable como la diferencia de talla y musculatura entre ambos sexos, aunque la división del trabajo entre varones cazadores y mujeres recolectoras haya dejado de condicionar nuestra evolución mental desde hace mucho tiempo.
Una hipótesis popularizada por libros y artículos de divulgación sugiere que los varones necesitaban una mayor capacidad de orientación porque la caza requiere explorar territorios más amplios que la recolección a poca distancia del campamento[141]. Una variante de esta idea cambia cazadores por guerreros: los varones se desplazaban por dominios más amplios que las mujeres en el contexto de la guerra intergrupal. En otro orden de cosas, una hipótesis alternativa apela a la selección sexual femenina. De acuerdo con esta idea, las mujeres habrían preferido como padres de sus hijos a los cazadores más competentes, y ser un buen cazador requiere aptitud espacial. Por último, incluso se ha sugerido que una aptitud espacial disminuida habría sido más una ventaja que un inconveniente para las hembras protohumanas, porque ello habría contribuido a que se mantuvieran cerca de casa, en particular durante el embarazo y la lactancia, y la consiguiente reducción de su movilidad habría incrementado su seguridad y les habría permitido ahorrar energía para la reproducción[142].
Soy más que escéptico hacia este estilo de adaptacionismo «panglossiano»[143], y más cuando la existencia misma del rasgo que se pretende explicar (en este caso un dimorfismo sexual psicológico) es altamente dudosa. Por descontado, no voy a cuestionar la relevancia de la división sexual del trabajo para la evolución humana (como sí hacen algunas antropólogas influidas por esa perversión filosófica llamada feminismo epistemológico). Dudar de que los machos homínidos cazaban y las hembras recolectaban, como ocurre en todas las sociedades preagrícolas que han llegado hasta nuestros días (una división sexual de la obtención del alimento que ya se insinúa en los chimpancés, los animales más cercanos a nosotros, y los más parecidos a nuestros ancestros protohomínidos), es anteponer el compromiso ideológico al buen juicio científico. Como ha señalado Marlene Zuk, bióloga y feminista militante, no se debe vetar la investigación y la comprensión biológica de las diferencias sexuales con el argumento de que ello contraviene el ideario feminista. No voy a negar la existencia de diferencias psicológicas innatas entre varones y mujeres, al menos en el ámbito restringido de la conducta sexual y reproductiva (como la orientación sexual, por poner un ejemplo obvio). Pero no veo por qué una aptitud mental ventajosa para un sexo no puede serlo también para el otro, aunque sea en un contexto diferente.
Topillos en el laberinto
Otro argumento esgrimido para justificar la mayor aptitud espacial masculina se basa en la idea de que los machos en general tienden a dominar territorios más extensos, porque en su búsqueda de parejas sexuales se desplazan por áreas que abarcan los territorios de varias hembras. Aquí la presión selectiva que habría propiciado el desarrollo de la aptitud espacial masculina habría sido la necesidad de desplazarse por un territorio amplio sin perderse. Obviamente, los animales no humanos no se orientan mediante mapas, pero se supone que facultades similares entran en juego cuando se trata de orientarse en un laberinto. Por eso las pruebas de habilidad para encontrar una recompensa o una salida en un laberinto son un recurso habitual de los investigadores para evaluar la aptitud espacial de una variedad de especies, sobre todo roedores. Pues bien, en algunas especies se ha detectado un dimorfismo sexual psicológico análogo al humano, al menos en ratas y algunos topillos (un grupo de roedores que incluye el hámster y el lemming).
El topillo campestre (Microtus pennsylvanicus) es una especie poligínica cuyos machos dominan un territorio que abarca los de varias hembras. Los topillos de las praderas (M. ochrogaster) y de los pinares (M. pinetorum), en cambio, son monógamos. Cuando se comparó la capacidad de desenvolverse en un laberinto de las tres especies, resultó que los topillos monógamos no mostraban dimorfismo sexual significativo en ningún caso, mientras que los machos de la especie poligínica encontraron su camino antes que las hembras a través de siete tipos distintos de laberinto[144]. Éste era un resultado esperable, porque cabe pensar que una aptitud espacial aumentada es una ventaja adaptativa para el sexo con mayor recorrido. También se notificó que las hembras con niveles bajos de estradiol (un estrógeno) lo hacían mejor que las hembras con niveles altos de la misma hormona, pero el otro resultado que me parece más relevante es que el mencionado dimorfismo sexual no se apreciaba en los topillos juveniles (a diferencia del caso humano, donde los niños comienzan a aventajar a las niñas en aptitud espacial desde edades tempranas). En consonancia con estos resultados, se ha reportado que el hipocampo, la parte del cerebro implicada en el razonamiento espacial, es mayor en los topillos machos poligínicos, mientras que los monógamos no exhiben dimorfismo cerebral[145].
Pero las cosas quizá no sean tan sencillas, como sugiere el caso del modelo animal favorito de los psicólogos experimentales, la rata de laboratorio. Algunos estudios habían establecido que también en esta especie los machos superaban a las hembras en aptitud espacial, pero investigaciones más recientes que abarcaban todo el ciclo ovulatorio femenino revelaron que los resultados de las hembras dependen de su estado reproductivo, mientras que los machos son más regulares. Si los autores de los estudios previos hubieran comparado la ejecución de ambos sexos diariamente a lo largo de varios ciclos ovulatorios, no habrían observado diferencias significativas (a lo sumo, sólo una ligera ventaja masculina en días concretos). Muchos científicos quizá considerarían que la diferencia es más relevante que la similitud, pero lo cierto es que el dimorfismo sexual es ínfimo y se esfuma cuando se comparan las medias masculina y femenina a lo largo de todo un ciclo reproductivo[146].
En cualquier caso, la masculinidad no tiene por qué implicar una mayor aptitud espacial o un hipocampo más desarrollado. Es cierto que esto parece ser así en los topillos poligínicos, pero no ocurre lo mismo en otros roedores. En el caso del ratón ciervo (Peromyscus maniculatus), otro roedor común en América del Norte, las hembras parecen desenvolverse mejor que los machos al menos en algunos tipos de laberinto (como el de Morris, una versión acuática donde los animales aprenden a encontrar una plataforma sumergida nadando en un tanque circular lleno de agua enturbiada mediante la adición de látex u otro material inerte). A veces las hembras tienen que resolver problemas espaciales más complejos que los machos. Un ejemplo bien estudiado es el del boyero (Molothrus ater), un ave parásita que pone sus huevos en los nidos de otras especies (el equivalente americano del cuclillo eurasiático). Las hembras de esta especie exploran grandes extensiones en busca de huéspedes potenciales, y tienen que volver a los nidos elegidos más de una vez, porque deben sincronizar el desarrollo de sus propios huevos con el de las puestas de sus huéspedes para asegurarse de que los polluelos parásitos rompan el cascarón a la vez que los otros, ya que de no ser así los padres putativos podrían rechazarlos. De esta manera, las hembras de boyero deben ser capaces de recordar la localización de cada nido parasitado para revisitarlos en el momento justo, lo cual requiere una aptitud espacial aumentada, por lo que no sorprende que posean un hipocampo más desarrollado que los machos[147].
Igualdad y diferencia
Lo que nos enseñan los dimorfismos sexuales cognitivos documentados en roedores y aves es que la historia natural de una especie puede dar sentido a diferencias neuropsicológicas favorables a uno u otro sexo. Ahora bien, ¿qué relevancia tienen las diferencias entre machos y hembras de topillos, ratones, canarios o boyeros para la probabilidad de que una mujer pueda llegar a ser una profesional competente en el campo de la ciencia o la ingeniería? A los psicólogos evolucionistas les gusta decir que varones y mujeres son casi como especies distintas. Pero la desigualdad sexual no debe darse por sentada. Las diferencias entre los sexos, cuando existen, son un problema evolutivo que requiere explicación (una explicación convincente, no un argumento simplista y ad hoc). Por mi parte, tengo claro que me parezco mucho más a una mujer que a un chimpancé, aunque sea macho (y, desde luego, mucho más que a un topillo). A veces esta declaración requiere más rotundidad de lo que podría pensarse, quizá porque nos empeñamos en encontrar y exagerar diferencias que sustenten nuestros sesgos ideológicos.
Puede que encontrar el camino de vuelta al campamento fuera una presión selectiva más intensa para los machos protohomínidos que para las hembras, o puede que no. En cualquier caso, debería tenerse muy en cuenta que la diferencia intersexual en los resultados de las pruebas de aptitud espacial y de aptitud matemática en general es apenas significativa. Las distribuciones estadísticas de las calificaciones de ambos sexos se solapan ampliamente (más aún que las estaturas, por ejemplo), así que los resultados de un test no permiten predecir el sexo de un individuo sin más. Es absurdo, por lo tanto, prejuzgar las expectativas profesionales de una persona a partir de su condición masculina o femenina. Las conclusiones de los estudios de diferencias psicológicas entre sexos no pretenden servir de justificación a los padres que motivan más a sus hijos que a sus hijas para sacar buenas notas en matemáticas y ciencias, o a los empleadores que rechazan a las mujeres por el mero hecho de serlo. Al menos en principio, quienes auspician y efectúan estos estudios lo hacen por su interés científico intrínseco (sea éste mucho o poco, lo cual es opinable). Los dimorfismos sexuales psicológicos pueden estudiarse y conocerse sin hacer juicios de valor, y sin exagerar su relevancia para nuestra vida diaria, lo que implica mantener la mente abierta y, a la vez, tener presente que nuestras preconcepciones pueden llevarnos a omitir ciertos datos y a aceptar acríticamente otros.
Por otra parte, la reducción de las diferencias entre sexos en las últimas décadas invita a pensar que la biología tiene bien poco que ver con la escasez de mujeres en campos como, por ejemplo, la ingeniería o las ciencias físicas. Es más, tan extendida como la creencia en la superioridad matemática masculina está la creencia en la superioridad verbal femenina, a pesar de lo cual nadie parece encontrar incongruente que los varones dominen los escalafones profesionales superiores de las humanidades tanto como los de las ciencias, y tampoco nadie haya insinuado que El Quijote fue escrito en realidad por una «negra» de Cervantes (aunque sí hay quienes sostienen que la teoría de la relatividad no fue concebida por Einstein, sino por su mujer). Dicho sea de paso, también se han propuesto explicaciones adaptacionistas de la presunta superioridad verbal femenina, como la que relaciona dicha aptitud con el prolongado vínculo maternofilial en nuestra especie[148]. La idea es que las madres mejor dotadas verbalmente estaban en mejor posición que los padres (que siempre andaban por ahí cazando) para enseñar a sus hijos pequeños a hablar. (Uno se pregunta por qué una facultad tan sofisticada debería haberse desarrollado más en las mujeres sólo para adiestrar linguísticamente a unos sujetos que balbucean palabras de una sílaba). En cualquier caso, la diferencia de aptitud verbal entre los sexos es aún menos significativa que la diferencia de aptitud matemática, y muchos psicólogos y educadores dudan de que sea real. Por mi parte, las afirmaciones del estilo de que las mujeres son más intuitivas y emotivas y tienen más capacidad verbal que los varones me parecen una reformulación políticamente correcta del viejo tópico sexista de que las mujeres son irracionales y parlanchinas.
La naturaleza es amoral y apolítica. Nuestra igualdad sexual es un resultado contingente de la evolución humana. La selección natural podría haber convertido a los machos homínidos en enanos descerebrados, como los machos de Ceratias. Pero no lo hizo, como tampoco les dotó de capacidades mentales ausentes o disminuidas en el otro sexo. En principio, no hay ninguna buena razón para pensar que la selección natural haya favorecido alguna diferencia intelectual innata entre varones y mujeres. Mientras no haya pruebas de lo contrario (pruebas fehacientes que vayan bastante más allá de los resultados de un test psicológico) es la igualdad sexual, y no la diferencia, la que debe darse por sentada.