
Figura 6.1. Un macho de pavo real pavoneándose, el icono de la selección sexual elegido por Darwin. (© Biopix.dk: N. Sloth).
La selección natural es el concepto central de la teoría darwiniana de la evolución, sintetizado a menudo en el famoso aforismo «La supervivencia del más apto» (que en realidad es de Spencer, no de Darwin, aunque éste lo suscribía al ciento por ciento). En la concepción darwiniana, la selección natural es básicamente una cuestión de aptitud para la supervivencia. De ahí que a Darwin le desconcertara sobremanera que, en muchas especies, los caracteres masculinos que atraen al otro sexo no sólo no facilitan en absoluto la supervivencia, sino que la comprometen. (Por ejemplo, los mismos ornamentos llamativos o cantos estridentes que atraen a las hembras también atraen a los predadores, y las colas aparatosas como la del pavo real o algunas aves del paraíso son un impedimento para el vuelo). Estos caracteres sexuales secundarios manifiestamente desventajosos parecían poner en tela de juicio la idea de que sólo los mejores supervivientes pasan el examen de la selección natural.
Para explicar los rasgos heredables que incrementan el éxito reproductivo a expensas de la aptitud para la supervivencia, Darwin se sacó de la manga la noción de «selección sexual». Éste es un concepto que siempre he encontrado un tanto superfluo, porque la selección natural no es una cuestión sólo de supervivencia, sino de eficacia biológica[109]. Lo que cuenta para la selección natural es la descendencia efectiva (es decir, no el número de hijos producidos, sino el de los que consiguen llegar a adultos y reproducirse a su vez). Por supuesto, para reproducirse es obligado estar vivo, pero la supervivencia estéril es irrelevante para la selección natural. En vez de «supervivencia del más apto», quizá sería más correcto hablar de «reproducción del más apto». Visto así, la selección sexual no es más que un aspecto particular de la selección natural, aunque pensar en términos de selección sexual pueda ser conveniente a la hora de considerar la coevolución de las biologías masculina y femenina.
Darwin distinguió dos modalidades de selección sexual, una ejercida por las hembras y otra ejercida por los propios machos. La primera, más propia de las aves, es la elección femenina de los machos más atractivos. El arquetipo elegido por Darwin para ilustrar este concepto, el pavo real, se ha convertido en un icono de la selección sexual (figura 6.1). Aunque su concepción de la selección natural se centraba en la capacidad de supervivencia, Darwin no podía creer que la aparatosa librea de los pavos reales les facilitara en algo la vida. Bien al contrario, la pesada cola es un impedimento a la hora de levantar el vuelo para escapar de los predadores. Pero el hecho de que los machos la desplegaran durante el cortejo llevó a Darwin a razonar que el motivo de tanta ostentación es que las hembras eligen como sementales a los machos de cola más imponente.
La segunda forma de selección sexual distinguida por Darwin no es otra que el combate físico entre los sementales por acaparar a las hembras. Si las hembras son un recurso limitante para la reproducción masculina, como suele ser el caso, los machos que se impongan a sus rivales en la lucha por el acceso a las hembras fecundables dejarán más descendencia. (En los casos de inversión de roles donde las hembras no ejercen de madres y el sexo masculino se encarga de la crianza, los machos pueden convertirse en un recurso limitante para la reproducción femenina, lo que propicia la evolución de hembras galanteadoras y combativas). Esta forma de selección puede propiciar la evolución de atributos costosos como cornamentas, colmillos o un tamaño corporal aumentado.
Figura 6.1. Un macho de pavo real pavoneándose, el icono de la selección sexual elegido por Darwin. (© Biopix.dk: N. Sloth).
Para Darwin, la selección sexual podía explicar la evolución de rasgos a los que no podía encontrárseles ningún valor adaptativo (en el sentido restringido de contribución a la supervivencia). Pero la propuesta fue recibida con escepticismo precisamente por sus correligionarios más fervientes. Lo que los darwinistas decimonónicos, más adaptacionistas que el propio Darwin, encontraban inaceptable era la arbitrariedad de la elección femenina. Wallace, por ejemplo, prefería la explicación alternativa de que eran las hembras, más necesitadas de ocultarse de los predadores, las que habían adquirido colores crípticos, mientras que los machos simplemente se habían mantenido en una condición prístina. En cambio, Wallace sí encontraba aceptable la otra modalidad de selección sexual, la derivada del combate físico entre los machos, quizá porque le parecía más acorde con la metáfora de la lucha por la vida. Del mismo parecer eran otros críticos que querían ver en las ostentaciones masculinas un modo de intimidar a los rivales (una idea desempolvada por algunos autores, cuya versión moderna se conoce como la hipótesis de la «propaganda de guerra»)[110]. Pero esta interpretación era difícil de conciliar con el hecho de que los pavos reales y otras aves desplegaran su cola ante las hembras y no lo hicieran ante los rivales.
En realidad, éste era un debate en buena medida espurio, porque, como acabo de recalcar, la selección sexual no es más que un aspecto particular de la selección natural. Y más importante aún que la lucha por la vida es la lucha por la reproducción. Las anatomías y comportamientos de los animales no tienen por qué haberse seleccionado siempre porque mejoran la capacidad de supervivencia, sino que pueden haberse adquirido sólo porque permiten más o mejores apareamientos. La selección natural adapta los individuos a su entorno, sí, pero en éste hay que incluir los competidores y los posibles socios reproductores. Así pues, la elección femenina no es más que otro factor limitante del éxito reproductivo masculino. Pero la idea de que un factor tan voluble pudiera llegar a producir «engendros» como el pavo real resultaba inaceptable para quienes, como Wallace, consideraban que la evolución no podía regirse por influencias caprichosas e insensatas, sino que debía conducir a soluciones óptimas.
Como hemos visto en el capítulo 3, hay buenas razones teóricas para esperar que las hembras seleccionen escrupulosamente a sus socios reproductores. Puesto que el progenitor que menos invierte en el negocio de la reproducción es también el que menos tiene que perder, se comprende que, por regla general, sean los machos los que se insinúan a las hembras y no al revés, y los que menos miramientos tienen a la hora de aparearse. Las hembras, en cambio, se juegan mucho más en el envite, lo que las obliga a evaluar de alguna manera la idoneidad del aspirante a padre de sus hijos. Pero esto es la teoría. En la práctica, a menudo la pretendida selectividad femenina es más virtual que otra cosa. Cuando la competencia por las hembras se resuelve a base de puños, los agentes de la selección sexual son los propios machos. Cualquier aspirante que haya sido capaz de imponerse a todos sus rivales ha acreditado sobradamente su aptitud, así que las hembras no necesitan ejercer una selección activa. La exclusión competitiva de los menos aptos, cuyo acceso a las hembras queda vedado, hace innecesaria la evolución de criterios de elección por parte femenina. (Hay que decir, no obstante, que incluso en este caso puede darse cierta discriminación sexual por parte femenina, pues no siempre los machos de rango máximo son la mejor elección, lo que es especialmente cierto cuando hay consanguinidad, en cuyo caso una hembra puede preferir a un macho extraño para evitar los costes del incesto). Sólo cuando esta modalidad de selección sexual es imperfecta o, por las razones que sean, los aspirantes no están por la labor de pelearse por las hembras, les toca a ellas elegir.
Hoy ya nadie pone en duda que el sexo femenino puede ejercer una selección sexual lo bastante efectiva para propiciar la exageración de los rasgos masculinos. Esto es fácilmente demostrable en aves como la viuda colilarga africana, cuyos machos poseen unas plumas caudales inusualmente largas. Pues bien, si se cortan las puntas de las colas de un grupo de machos y los segmentos de pluma cortados se emplean para alargar artificialmente las colas de otro grupo de machos, se observa que estos últimos se aparean más veces que los machos de cola normal, y éstos más que los de cola recortada. El mismo criterio de elección femenina parece regir en muchas otras aves con un dimorfismo sexual semejante[111].
Darwin tenía razón. Pero la controversia decimonónica sobre la selección sexual no tenía tanto que ver con su realidad como con su sentido adaptativo. Más que las colas vistosas mismas, era la preferencia femenina por las colas vistosas lo que requería una explicación satisfactoria (es decir, adaptacionista). Este debate nunca se zanjó del todo, y ha vuelto a reavivarse un siglo después. En un bando están los que afirman que las hembras eligen machos atractivos porque quieren tener hijos igual de atractivos que puedan darles muchos nietos portadores de sus genes. Es lo que se conoce informalmente como la hipótesis del «hijo sexy»[112]. En el otro bando están los que encuentran insatisfactoria esta explicación e insisten en que la elección femenina de los apareamientos debe basarse en criterios intrínsecamente relacionados con la calidad genética real del aspirante a padre. Los primeros convienen con Darwin en que la elección femenina no tiene por qué regirse por criterios «objetivos» de aptitud darwiniana, sino que puede obedecer a criterios estéticos arbitrarios. Los segundos comparten el rechazo de Wallace hacia la idea de una selección sexual esencialmente arbitraria, y replican que la preferencia femenina por los ornamentos ostentosos o los cantos sonoros debe ser adaptativa en sí misma.
Ronald Fisher ya indicó que la preferencia de una hembra por las colas largas o cualquier otro adorno arbitrario se justifica por sí sola si las otras hembras comparten la misma preferencia[113]. Si el grueso de la población femenina prefiere los machos de cola larga, entonces una hembra que se deje fecundar por un macho de cola corta estará comprometiendo el futuro de sus genes, porque si sus descendientes masculinos heredan la falta de atractivo del padre tendrán escaso éxito reproductivo y, en consecuencia, serán pésimos propagadores de los genes maternos. Así pues, el mejor partido para cualquier hembra sería el tipo masculino más atractivo para la mayoría. Pero este argumento sólo justifica por qué se mantiene un criterio de apareamiento determinado, como el de las colas largas, no por qué se selecciona en primera instancia hasta hacerse mayoritario. Una respuesta apuntada por el propio Fisher es que el criterio en cuestión no sea del todo arbitrario de entrada, sino que tenga alguna relación con la aptitud darwiniana. Así, el tamaño y aspecto de la cola podría ser un indicador de la edad y la condición física del pretendiente: una cola comparativamente corta y deslucida puede ser indicativa de subdesarrollo por enfermedad, debilidad constitucional o malnutrición derivada de ineptitud para obtener alimento o competir por él, mientras que una cola lustrosa y bien desarrollada puede ser un estimador razonablemente fiable de la aptitud de su dueño. De hecho, son legión las especies de aves cuyo cortejo incluye el despliegue de la cola, aunque ésta no sea un ornamento en sí misma, lo que sugiere que las hembras evalúan la idoneidad del pretendiente a partir del aspecto de su plumaje.
Una hembra que elija sus parejas sexuales según un criterio que tenga que ver con la aptitud darwiniana de los padres de sus hijos dejará más y mejor descendencia, con lo que el criterio de elección (que suponemos innato y heredable) se seleccionará junto con su progenie, y una vez las hembras que lo hayan heredado constituyan una fracción lo bastante significativa de la población, la selección sexual hará que el criterio de elección se justifique por sí mismo, con independencia de su sentido adaptativo inicial. En esta línea de pensamiento, se ha sugerido que las libreas vistosas son indicadores de inmunidad a los parásitos de toda clase, desde las lombrices hasta los virus[114]. Esta propuesta se basa en la observación de que las especies más vistosas suelen ser también las más hostigadas por los parásitos[115]. Claro que esta correlación podría ser acausal, porque tanto los parásitos como las coloraciones llamativas abundan en los ecosistemas tropicales. Pero diversos estudios han confirmado que los machos con menor carga de parásitos suelen ser también los elegidos con preferencia por las hembras. Esto parece valer para una amplia variedad de especies, desde los faisanes hasta los grillos. Un ejemplo bien estudiado es el de las golondrinas, cuyas hembras tienen preferencia por los machos de cola más larga y simétrica. Pues bien, parece ser que los machos de cola más simétrica también son los menos afectados por los ácaros[116]. En el caso del gallo de las artemisas, un pariente norteamericano del urogallo, la conexión entre el atractivo masculino y la carga de parásitos no puede ser más directa. Los machos de esta especie se congregan en una palestra (lo que los biólogos llaman «lek») y cada uno defiende un espacio propio donde se dedica a exhibirse hinchando dos sacos pectorales de color amarillo vivo. Los gallos más viejos tienden a ocupar las posiciones centrales del área de exhibición, y suelen ser los preferidos por las hembras que acuden a aparearse. A menos que los sacos pectorales delaten picaduras de piojos: basta con pintar puntos rojos en los sacos aéreos de un macho de aspecto por lo demás saludable para que su atractivo sexual caiga en picado, aunque ocupe el centro de la palestra[117].
Sea como fuere, lo cierto es que los modelos teóricos no cierran la puerta a la posibilidad de que la selección sexual pueda partir de cero, sin necesidad de que la preferencia femenina tenga valor adaptativo alguno[118]. Si se parte de una condición inicial tal que todos los machos sean igualmente aptos y los apareamientos completamente al azar, entonces cualquier preferencia que se introduzca, por arbitraria que sea, perturbará la distribución uniforme de la eficacia biológica masculina. Puesto que el rasgo masculino atrayente y la atracción femenina por el mismo se refuerzan mutuamente, esta desviación de la uniformidad estadística, aunque sea mínima, bastará para iniciar un bucle retroactivo positivo que llevará a la evolución de rasgos físicos y/o conductuales cada vez más ostentosos. Este proceso de selección sexual «desbocada» (como lo describió Ronald Fisher) sólo se frenará cuando la exageración de los caracteres masculinos se convierta en una carga tan pesada que la ganancia de atractivo sexual no compense el coste para la supervivencia.
La uniformidad de la aptitud masculina no es una condición inicial tan poco realista como puede parecer a primera vista, porque, como hemos visto en el capítulo 3, en un sistema de apareamiento poligínico la selección continuada de los mejores entre los mejores conduce a una igualdad creciente entre los aspirantes a padre, de manera que una hembra siempre tendrá a su disposición pretendientes de sobra que satisfagan todos los requerimientos de calidad genética. En estas condiciones parece más factible la evolución de criterios de apareamiento arbitrarios. El hecho de que sea precisamente en las especies altamente poligínicas, como el pavo real o el faisán dorado, donde el barroquismo de las libreas masculinas se haga más extremado viene a confirmar esta predicción.
Otro aspecto poco realista de los modelos teóricos es la omisión del coste de la elección femenina; y es que discriminar entre los pretendientes requiere una inversión no nula de tiempo y energía. Pues bien, si se introduce este coste, aunque sea ínfimo, en los modelos teóricos, resulta que ninguna preferencia absolutamente arbitraria puede prosperar. Aunque la selección sexual desbocada pueda favorecer inicialmente la difusión de una preferencia femenina caprichosa, una vez la exageración del rasgo masculino atrayente se convierte en una carga demasiado pesada, la selección natural vuelve a favorecer a las hembras no selectivas frente a las que continúan expresando la preferencia en cuestión[119]. Salvo en el caso de que exista un sesgo mutacional. En otras palabras, si es más probable que una mutación aleatoria acarree una pérdida de atractivo que una ganancia, entonces para las hembras continuará siendo rentable pagar el coste de la elección de pareja sexual. Al final se llega a un compromiso tal que el coste de la elección queda compensado por la ventaja de tener hijos más atractivos que la media[120]. Dicho sesgo mutacional es tanto más esperable cuanto más elaboradas y extremadas sean las libreas masculinas. También es de esperar un sesgo mutacional en la viabilidad, pues cuanto mayor es la complejidad de un sistema vivo y más óptima es su adaptación al entorno físico y biótico, más probable es que una mutación aleatoria reduzca su aptitud darwiniana en vez de aumentarla. En la práctica, ambos sesgos mutacionales tenderán a ir de la mano, por lo que cabe esperar que los criterios femeninos de belleza masculina estén ligados (al menos en parte, y aunque sea de manera muy indirecta) a la aptitud de los pretendientes.
Por las razones expuestas, una selección sexual basada en criterios absolutamente arbitrarios no parece verosímil fuera de los modelos matemáticos. No en vano la razón última de la selectividad femenina es aparearse con machos de calidad genética contrastada. De hecho, el aporte de genes frescos es la principal razón de ser del sexo masculino (al menos en las especies en las que el sexo femenino asume la totalidad de la inversión parental). Es de suponer, por lo tanto, que la selección sexual ejercida por las hembras obedece a criterios no arbitrarios, aunque a caballo de éstos puedan evolucionar preferencias gratuitas (sobre todo en las especies altamente poligínicas). Numerosas observaciones y experimentos de campo abonan la tesis de que los machos preferidos por las hembras como sementales son también los más curtidos, vigorosos y saludables. Ahora bien, si se acepta que los criterios de elección femeninos están encaminados a evaluar la calidad genética de los aspirantes a padre, ¿por qué la selección sexual ha promovido la evolución de armas de seducción tan costosas para sus dueños como la cola del pavo real? Si se trata de elegir a los machos mejor dotados para la supervivencia, ¿por qué dejarse encandilar por unos atributos masculinos desmesurados que comprometen la supervivencia de su poseedor?
El israelí Amotz Zahavi ofreció una controvertida solución a esta paradoja. La idea es que, cuanto más costoso es un modo de ostentación masculina, mayor es su fiabilidad como indicador de aptitud darwiniana. Todo macho que consigue salir adelante aun cargando con un lastre innecesario está enviando el mensaje de que es tan superior a sus competidores que puede permitirse jugar con desventaja. La implicación es que un criterio de apareamiento basado en un atributo genuinamente desventajoso (y, por ende, fiable) será favorecido por la selección natural frente a cualquier preferencia femenina arbitraria[121].
Como ocurre a menudo con las buenas ideas científicas, la conjetura de Zahavi, conocida como «principio del hándicap», fue recibida con escepticismo. Dawkins llegó a ridiculizarla con el contraargumento de que un corolario lógico de dicho principio sería la evolución de machos cojos y tuertos de nacimiento. Para otros, la idea tiene un tufillo tautológico (una sospecha a la cual no ha dejado de contribuir el propio Zahavi, quien en su libro The Handicap Principle: A Missing Piece of Darwin’s Puzzle estira su principio hasta convertirlo en una suerte de teoría de casi todo, desde las castas obreras de los insectos sociales hasta la actividad sexual superflua de las parejas humanas o la evolución del lenguaje)[122]. Pero la principal objeción de los críticos era que el coste del hándicap anularía el beneficio de su fiabilidad como indicador de aptitud, porque la descendencia de una hembra que aplicase un criterio de elección semejante heredaría tanto los genes óptimos como el hándicap del padre. Ahora bien, si el hándicap se asocia a un rasgo ligado al sexo (en vez de uno neutro como la posesión de un par de ojos o un par de piernas), entonces las hijas heredarán los genes óptimos del padre pero no su hándicap. En cuanto a los hijos, heredar el hándicap paterno (y, por ende, el atractivo sexual aumentado) puede beneficiarles más que perjudicarles.
Al final, las reticencias iniciales han dado paso a la aceptación mayoritaria (hasta el propio Dawkins ha reconocido que Zahavi podría tener algo de razón). En el dominio de la selección sexual, el principio del hándicap ofrece una explicación adaptacionista satisfactoria de la exageración de los caracteres sexuales secundarios en los machos poligínicos cuyo éxito reproductivo depende sobre todo de la elección femenina. Es más, un corolario del mismo principio es que cualquier rasgo ligado al sexo puede valer como criterio de elección de pareja sexual si, como consecuencia de un proceso de selección desbocada, acaba convirtiéndose en un hándicap. Aun suponiendo que la evolución de la ostentosa cola del pavo real hubiera partido de un criterio de elección puramente estético, vacío de contenido adaptativo, la exageración creciente del rasgo habría convertido una preferencia inicialmente arbitraria en un criterio fiable de evaluación de la aptitud del semental de turno. Así pues, la selección sexual desbocada de Fisher y el principio del hándicap de Zahavi no sólo no son contradictorios, sino que se complementan a la hora de explicar la evolución de rasgos masculinos aparentemente paradójicos.
Quienes se resisten a contemplar la reproducción sexual como una empresa cooperativa y prefieren verla como una suerte de explotación mutua (más de las hembras por los machos que al revés) gustan de comparar las exhibiciones de cortejo con la publicidad. Hay un anunciante (en este caso el macho) cuyo principal interés es vender su producto (en este caso sus propios genes) que intenta engatusar a un comprador potencial (en este caso la hembra) cuyo principal interés es que no le den gato por liebre. Obviamente, la función de la publicidad no es dar información fidedigna sobre el producto, sino seducir a los compradores potenciales, aunque para ello tenga que falsear la realidad. Si el éxito reproductivo masculino depende de la elección femenina, sólo la fachada puede salvar a un macho mediocre de la muerte genética. Las hembras deben evitar dejarse engañar, para lo cual deben seleccionar sus apareamientos aplicando criterios que excluyan la publicidad engañosa.
Aquí es donde vuelve a entrar en escena el principio del hándicap, porque cuanto más costosos sean los alardes masculinos, más difícil será falsearlos. La preferencia femenina por los atributos costosos se justifica porque éstos suponen una carga tanto más pesada cuanto peor dotado está un macho para la supervivencia y la competencia con los rivales. Así pues, cualquier hándicap genuino es una forma de publicidad honesta (valga la contradicción), en el sentido de que proporciona información fidedigna acerca de la aptitud del macho que hace ostentación del mismo[123].
Por supuesto, el concepto «honestidad» resulta difícil de digerir para los adeptos a la metáfora del egoísmo genético, quienes insisten en que el interés primordial de todo macho no es acreditar su aptitud real ante sus «compradoras» potenciales, sino engatusar a cuantas más mejor. Ahora bien, desde el punto de vista estrictamente darwinista, el interés primordial de todo macho no es seducir a cuantas más hembras mejor, sino propagar sus genes más que sus rivales. Una hembra no es una «compradora», sino un socio reproductor al que el macho debe convencer para que invierta su capital en la empresa común de perpetuar los genes de ambos. Si uno puede ofrecer lo que el otro está buscando, entonces la mejor propaganda es una demostración no falseable de la propia valía. Esto se aplica especialmente a los regímenes poligínicos, porque cuanto más intensa es la selección sexual y mayor la igualdad entre los aspirantes a padre, más cuesta destacar sobre los rivales para acaparar la atención femenina. La competencia masculina exacerbada puede explicar el hecho frecuente de que la exhibición de libreas llamativas durante el cortejo vaya acompañada de sonidos estridentes, brincos y otros alardes de poderío físico difícilmente falseables.
Esto nos devuelve a la otra modalidad de selección sexual distinguida por Darwin: el combate físico. De hecho, la frontera entre ambos modos de selección sexual es borrosa. En la gran mayoría de casos, el éxito reproductivo masculino depende tanto de la elección femenina como de la exclusión de los rivales. Por eso no es infrecuente que una misma estructura o conducta tenga la función doble de disuadir a los rivales y seducir a las hembras. (Un ejemplo es la cornamenta de los ciervos, que además de servir para el combate por el dominio de los territorios frecuentados por las hembras es un indicador de la edad y la condición física de su poseedor). Una variación sobre este tema, la hipótesis de la «propaganda de guerra», atribuye esta función doble no sólo a los rasgos anatómicos, sino a los rituales de cortejo[124]. La idea es que los gritos, saltos y otros derroches de energía que suelen acompañar la exhibición de las libreas masculinas no sólo sirven para impresionar a las hembras, sino para intimidar a los machos rivales. Un ejemplo ilustrativo de esta dualidad es el caso del grillo maya (Amphiacusta maya). Como hacen todos los grillos machos, los de esta especie cantan para atraer a las hembras fecundables, pero al hacerlo atraen también a los rivales, formándose congregaciones de machos que cantan al unísono (lo que incrementa sobremanera el volumen y alcance de su mensaje sonoro) y cuyo acceso a las hembras se rige por una estricta jerarquía de dominancia. Pues bien, si se embadurnan los élitros de los machos más dominantes con cera para amortiguar su canto, se observa que sus rivales los desafían sin tregua. Los machos así tratados suelen acabar imponiéndose igualmente a sus retadores, lo que indica que su superioridad es genuina, pero la incapacidad de emitir un canto disuasorio les obliga a malgastar tiempo y energía en peleas evitables, en detrimento del número de apareamientos[125]. Aunque todo macho tiene que pagar una cuota por acceder al rango más alto, resolver las disputas cantando nunca resulta tan costoso como hacerlo a base de puños.
Por las razones expuestas, es de esperar que los mensajes destinados al otro sexo sean «publicidad honesta». Por supuesto, a los individuos menos aptos siempre les convendrá que tanto los socios reproductores como los rivales se dejen embaucar. La fiabilidad de los indicadores de aptitud es producto de la competencia entre los candidatos a socio reproductor en un régimen de selección sexual. Son los mejor dotados los que deben adoptar indicadores de calidad genética no falseables, y sus posibles socios reproductores los que deben protegerse de la publicidad engañosa aplicando criterios de elección no arbitrarios. La comunicación honesta entre los sexos es el resultado final de un doble proceso de selección en virtud del cual las señales falseables o insustanciales acaban siendo devaluadas por los rivales e ignoradas por las hembras.
Cuando los machos aportan algo más que sus genes y la inversión paterna en la crianza se convierte en un recurso valioso, la elección del mejor semental posible debe subordinarse a la conveniencia de contar con un consorte proveedor. Una hembra monógama cuyas exigencias le hagan demorar en exceso su emparejamiento será penalizada por la selección natural, pues los machos más cotizados suelen ser los primeros en emparejarse, y no siempre se llega a tiempo de adelantarse a las rivales. Si la contribución masculina es decisiva para una crianza exitosa, un consorte mediocre es mejor que nada. Así pues, los criterios de elección de pareja en un régimen monógamo deben ser más flexibles que en un régimen poligínico. Esto no significa que las hembras monógamas concedan menos importancia al atractivo masculino que las autosuficientes. Cuando las hembras sólo buscan sementales y no padres de familia, la selección sexual es tan intensa que la oferta de calidad genética masculina supera con creces la demanda, así que una hembra puede permitirse rechazar a cualquier candidato que no le parezca absolutamente perfecto, porque siempre habrá otros que satisfagan sus exigencias. En un sistema de apareamiento monógamo, en cambio, la demanda de calidad genética masculina siempre es mayor que la oferta, pues la aptitud de los candidatos es mucho más variable y los mejores son los primeros que dejan de estar disponibles. Las hembras ya no pueden limitarse a rechazar a cualquier macho que no juzguen perfecto, sino que deben evaluar la calidad genética de los machos disponibles para elegir al mejor partido (o el menos malo) si quieren rentabilizar al máximo su inversión parental. Esto implica que los criterios de elección de pareja aplicados por las hembras monógamas deben ser todavía menos arbitrarios que los aplicados por las hembras autosuficientes.
En cualquier caso, la distribución del éxito reproductivo masculino en un régimen monógamo es mucho más uniforme que en un régimen poligínico, lo que explica la práctica ausencia de dimorfismo sexual (es decir, la diferencia de tamaño y apariencia entre machos y hembras) en las especies monógamas típicas. Incluso los ornitólogos avezados tienen serias dificultades para identificar el sexo de cada miembro de una pareja de albatros, pingüinos o cigüeñas. Aun así, se sabe que las hembras monógamas también aplican criterios de elección de pareja basados en rasgos físicos. Por ejemplo, se ha comprobado que los machos de golondrina a los que se alarga artificialmente la cola se emparejan antes y efectúan más cópulas extraconyugales que los no manipulados[126]. Esta selección sexual puede explicar la existencia de rasgos dimórficos en muchas especies nominalmente monógamas, pero facultativamente polígamas o proclives al sexo extraconyugal, aunque nunca tan exagerados como los resultados de la selección sexual desbocada. Este dimorfismo sexual moderado es bastante corriente en las aves migratorias con las que estamos familiarizados. Pero ya hemos visto que casi todas estas aves nominalmente monógamas son en realidad facultativamente polígamas y proclives al adulterio. En la mayoría de casos las parejas se deshacen al concluir la temporada de cría, y cada año se repite el ritual de cortejo y emparejamiento. Los machos suelen adelantar su migración primaveral para tomar posesión de un buen territorio de nidificación, así que las primeras hembras en llegar tienen pretendientes de sobra para elegir. Un macho atractivo no sólo puede emparejarse sin pérdida de tiempo, sino que tiene más posibilidades de vincularse a una hembra secundaria, lo que le permite sacar adelante más de una nidada por temporada. Por otra parte, las hembras ya emparejadas suelen estar dispuestas a dejarse inseminar por un macho más seductor que su consorte si tienen ocasión. La elección femenina contribuye así a que unos machos dejen más descendencia que otros, condición necesaria para que se dé una selección sexual lo bastante significativa para propiciar la evolución de caracteres masculinos secundarios.
Una tercera forma de selección sexual, más sutil que los concursos de belleza o el combate físico, es la competencia espermática. Allí donde las hembras se aparean con más de un macho, éstos pueden adquirir caracteres que les ayuden a vencer en la carrera hacia el óvulo, como por ejemplo unos testículos capaces de producir espermatozoides supernumerarios. Un ejemplo instructivo y muy cercano de correlación entre promiscuidad femenina y tamaño testicular es el de los hominoideos. Los gorilas pesan cuatro veces más que los chimpancés, pero éstos tienen unos testículos cuatro veces mayores. Puesto que las gorilas sólo se aparean con el dueño de su harén, un gorila macho no necesita producir demasiados espermatozoides para asegurar la fecundación del eventual óvulo. Las chimpancés, en cambio, se aparean con varios machos, lo que suscita la competencia espermática y, por ende, el agrandamiento de los testículos. Esto vale también para el resto de primates: a más competencia espermática, mayor tamaño testicular, y quien se lleva la palma es el mono araña lanudo, cuyas hembras copulan con todos y cada uno de los machos de su tropa cuando están en celo, lo que ha propiciado que los machos de esta especie posean los testículos más grandes de todos los primates.
Una correlación similar se ha observado en otros grupos de mamíferos y en las aves. En estas últimas, los testículos más grandes pertenecen a los machos de especies poliándricas cuyas hembras compiten por harenes de machos, como es el caso de las jacanas, mientras que los testículos más pequeños se encuentran en los regímenes poligínicos de lek, donde unos pocos machos elegidos acaparan los apareamientos y la competencia espermática es prácticamente nula, porque las hembras raramente se aparean con más de un semental.
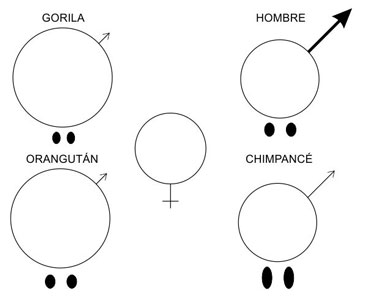
Figura 6.2. Dimorfismo sexual y competencia espermática en los machos hominoideos. En gorilas y orangutanes la selección sexual se basa en la fuerza bruta, mientras que en los chimpancés se basa más en la competencia espermática, lo que ha propiciado el agrandamiento de los testículos. En el caso humano la hipertrofia testicular no pasa de moderada, y es el pene el que ha experimentado un agrandamiento más significativo. (Modificado de Short, 1979).
Hay que hacer notar que los resultados evolutivos esperables de la competencia espermática poscopulatoria son diferentes de los resultados de la selección sexual precopulatoria. Por ejemplo, es presumible que la superioridad en el combate físico sea más decisiva en las especies poligínicas que en las monógamas, pero cuando las hembras se aparean con más de un macho también es de esperar un agrandamiento de los testículos. La pauta esperable es que los machos monógamos posean unos testículos comparativamente pequeños y exhiban un dimorfismo sexual ínfimo (aunque, como acabamos de ver, la infidelidad femenina puede convertir la monogamia en una poliginia efectiva capaz de suscitar una competencia precopulatoria y poscopulatoria lo bastante significativa para propiciar tanto el dimorfismo de tamaño como la hipertrofia testicular, aunque nunca tanto como en las especies genuinamente poligínicas), mientras que los machos que compiten por el acceso exclusivo a numerosas hembras que sólo se dejan inseminar una vez no deberían necesitar unos testículos grandes, pero sí un mayor tamaño corporal, quizá suplementado con armas como unos cuernos o unos caninos amenazadores; por último, los machos de especies poligínicas o poliándricas cuyas hembras son más o menos promiscuas pueden combinar el dimorfismo sexual con la hipertrofia testicular (como ocurre en los chimpancés). Así pues, se puede predecir el grado de poliginia y promiscuidad de una especie sin más que examinar el dimorfismo sexual y el desarrollo de los testículos.
La existencia de criterios masculinos de belleza femenina parece difícil de conciliar con la tesis sociobiológica clásica de que los machos son poco o nada selectivos a la hora de aparearse. Pero lo cierto es que ésta no es una ley biológica universal, ni mucho menos. Como hemos visto en los capítulos 3 y 4, el precio de la reproducción no es menor para el sexo masculino que para el femenino, porque los machos que se ahorran la inversión parental deben invertir un esfuerzo aún mayor en la competencia por los apareamientos. Por otra parte, para un macho no siempre es factible impedir que algún rival consiga acceder a las hembras de su dominio a sus espaldas. De ahí que incluso los machos poligínicos tiendan a concentrar su celo en ciertas hembras favoritas. Así ocurre, por ejemplo, en los chimpancés, los papiones y otros primates sociales cuyos machos dominantes prestan atención preferente a unas cuantas hembras favoritas en lugar de intentar en vano acapararlas a todas. Los machos de estas especies pueden preferir aparearse con hembras de alto rango, cuya prole tiene más posibilidades de salir adelante, o con madres expertas, cuya fecundidad está más garantizada. Aun así, en general sí puede afirmarse que los machos atienden más a la cantidad que a la calidad de sus apareamientos.
Ahora bien, si la inversión paterna en la crianza es comparable a la materna, o mayor, entonces los machos no tienen por qué ser menos selectivos que las hembras a la hora de emparejarse. Si la monogamia obliga a las hembras a flexibilizar sus criterios de elección para acomodarlos a la oferta masculina disponible, a los machos les obliga a tener en cuenta la fertilidad potencial y la aptitud de las candidatas a consorte. Y si incluso en un sistema de apareamiento monógamo las hembras pueden suscitar la evolución de caracteres masculinos secundarios, cabe preguntarse si los machos pueden ejercer una selección sexual lo bastante significativa para condicionar la evolución de caracteres femeninos secundarios. Esto resulta obvio en las jacanas y otras aves poliándricas cuyos machos ejercen de «madres», lo que suscita una selección sexual (atribuible tanto a la elección masculina como a la competencia femenina por los machos disponibles) que ha producido hembras más grandes y de plumaje más vistoso que los machos. Pero, salvo estos casos de inversión de roles, en general no parece que las preferencias masculinas susciten la evolución de caracteres femeninos exagerados.
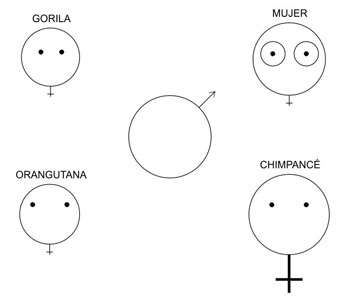
Figura 6.3. Las hembras hominoideas vistas por los machos. La hembra humana es la única que ha adquirido caracteres sexuales secundarios, en particular sus mamas rellenas de tejido adiposo, cuya «hinchazón» es permanente, a diferencia de las hinchazones genitales transitorias de la hembra chimpancé, que se ciñen a la fase fértil del ciclo menstrual. (Modificado de Short, 1979).
Hay, sin embargo, una notoria excepción, y no es otra que la hembra humana (figura 6.3). Algunos autores sostienen que la peculiar anatomía femenina humana se explicaría, al menos en parte, por un proceso de selección sexual fisheriana invertida (es decir, con las hembras como objetos de selección y los machos como sujetos). La idea es que una preferencia masculina inicialmente adaptativa por las mamas bien desarrolladas y las caderas anchas (signos obvios de aptitud reproductiva) habría propiciado la evolución de las adiposidades en mamas y caderas que proporcionan al cuerpo de la hembra humana su silueta característica. Pero está claro que el tejido adiposo no produce leche ni ensancha el canal del parto, por lo que algunos han querido interpretar las curvas femeninas como una forma de publicidad engañosa. Para embaucar a los machos homínidos y enviarles información falsa sobre el volumen de sus glándulas mamarias y la anchura de sus caderas, las hembras no habrían tenido más que engordar. Como contrapartida, la selección natural habría favorecido a los varones que se decantaran por las delgadas, lo que garantizaba la autenticidad de las mamas y caderas indicadoras de aptitud reproductiva. Las embaucadoras, por su parte, habrían recurrido al truco de mantener la cintura estrecha sin dejar de acumular grasa en mamas y caderas, con lo que se habría iniciado una selección sexual desbocada que habría exagerado unos indicadores de fecundidad potencial inicialmente honestos, hasta que los indicadores falseados se habrían mantenido simplemente por el atractivo que ejercían sobre los varones[127].
Pero este argumento, que no es más que el recíproco de la hipótesis del «hijo sexy», no tiene en cuenta que la selección sexual nunca actúa de manera simétrica. Recordemos que el potencial reproductivo femenino es muy inferior al masculino, lo que reduce drásticamente las posibilidades evolutivas de una exageración ficticia de los rasgos femeninos. Un hijo atractivo siempre será un propagador eficiente de los genes maternos, pues incluso en un régimen monógamo podrá engendrar descendencia extra a través del sexo extraconyugal o la poligamia facultativa, lo que puede suscitar una selección sexual capaz de originar un dimorfismo sexual apreciable. Pero no resulta tan obvio que una hija atractiva sea mucho mejor propagadora de los genes paternos que una hija poco agraciada, pues el éxito reproductivo femenino no depende (por lo menos no directamente) del número de parejas sexuales. Esto implica que el éxito reproductivo femenino está mucho menos ligado al atractivo sexual que el masculino. En una situación de competencia por los mejores partidos, la principal ventaja de ser más atractiva que las hembras rivales sería tener más posibilidades de emparejarse con un macho cotizado. Recordemos que la ventaja selectiva de tener hijos de un padre atractivo (suponiendo que los hijos tiendan a heredar el éxito reproductivo paterno) se hace efectiva al cabo de dos generaciones, en la forma de más nietos (cada uno de los cuales es portador de al menos una cuarta parte de los genes de su abuela). De esta manera, los genes responsables de los caracteres secundarios femeninos podrían propagarse a caballo del potencial reproductivo masculino.
A un macho, en cambio, emparejarse con una hembra atractiva que le diera hijas atractivas no le reportaría muchos más nietos, aunque quizá sí más bisnietos (cada uno de los cuales sería portador de al menos una octava parte de los genes de su bisabuelo) si las hijas atractivas lo tuvieran más fácil para unirse a yernos atractivos que le dieran nietos atractivos que fueran propagadores eficientes de los genes del abuelo. Pero ésta parece una ventaja bastante exigua en un régimen monógamo, donde el éxito reproductivo se reparte mucho más uniformemente que en un régimen poligínico (donde los machos no necesitan seleccionar sus apareamientos en función del atractivo femenino). Así pues, no parece demasiado razonable pensar que los caracteres secundarios femeninos son producto de una selección sexual fisheriana, y menos aún que no son más que «señuelos» para engatusar al sexo masculino. Bien al contrario, difícilmente se habrían seleccionado si no fueran indicadores honestos y fidedignos de aptitud y fecundidad potencial[128].