2
Sexo, parásitos e incertidumbre
Las hipótesis sobre la prevalencia de la reproducción sexual basadas en la diversificación de la descendencia sólo se tienen en pie si diferenciarse de los progenitores constituye una ventaja selectiva. En otras palabras, si lo que es más apto para una generación deja de serlo para la siguiente, lo cual es tanto más cierto cuanto más largo es el tiempo de generación (y, por ende, más probable es que las cosas hayan cambiado de una generación a otra). Como acabamos de ver, las observaciones parecen contradecir la idea de que la creación sexual de variantes es un requisito obligado para la persistencia y evolución de cualquier forma de vida en un entorno cambiante, porque las formas de vida asexuales medran en los ambientes fluctuantes. Pero las premisas de las que se deriva esta conclusión quizá sean demasiado simplistas.
Aunque los evolucionistas siempre han tendido a pensar más en términos de cambios físicos que de cambios en el entorno biótico de los organismos, lo cierto es que el fracaso o éxito reproductivo de un individuo pocas veces depende primariamente de factores no bióticos. Pensemos, por ejemplo, en la coevolución entre depredadores y presas. Las gacelas deben correr más deprisa que los guepardos para evitar ser devoradas, y los guepardos deben correr más deprisa que las gacelas para atraparlas. Se entabla así una «carrera de armamentos» evolutiva en la que depredadores y presas se hacen cada vez más veloces. Como la Reina Roja de Lewis Carroll en Alicia a través del espejo, los actores del drama evolutivo tienen que correr sin parar sólo para mantenerse en el mismo sitio. Además de los macrodepredadores, la presión de los competidores y, sobre todo, los parásitos de toda clase suelen ser factores más determinantes del éxito reproductivo que cualquier variable física[27].
Sexo contra parásitos
Los organismos deben adaptarse a un entorno biótico en constante evolución. Esto vale especialmente para los parásitos, que ejercen una presión selectiva más intensa que la de cualquier macrodepredador. Para empezar, son mucho más numerosos. Los conejos, por ejemplo, son presa de rapaces, mustélidos y zorros (además de perros y personas), bastante poco en comparación con la hueste de insectos, ácaros, gusanos, protozoos, bacterias, hongos y virus que los infestan. (El virus de la mixomatosis, por ejemplo, mata muchos más conejos que los zorros o cualquier otro macrodepredador natural). Al igual que depredadores y presas, parásitos y huéspedes están enlazados en un bucle retroactivo evolutivo. Cuanto más eficaz es el parásito (cuantos más huéspedes infecta o más recursos expolia) la selección natural más favorecerá a los huéspedes capaces de inventar una defensa; y cuanto mejores sean las defensas del huésped, la selección natural más favorecerá a los parásitos capaces de vencerlas o burlarlas. Pero lo más relevante aquí es que los parásitos suelen tener un ciclo vital mucho más corto que sus huéspedes, lo que quiere decir que por cada generación del huésped pueden sucederse muchas generaciones de parásitos. En el lapso de una generación del huésped, los parásitos de ciclo vital más breve están en condiciones de evolucionar y «aprender» a vencer unas defensas hasta entonces efectivas, de manera que un genotipo que antes confería resistencia puede quedar obsoleto de una generación a otra. Así pues, los parásitos pueden proporcionar el incentivo para diversificar los genotipos filiales que parece requerir la prevalencia del sexo.
Para abrirse paso hasta el interior de las células del huésped, los microbios y virus se fijan a la superficie celular mediante proteínas que encajan como llave y cerradura. Los parásitos encuentran la llave que abre la cerradura, los huéspedes cambian la cerradura, y así sucesivamente. De nuevo, la ventaja del sexo puede justificarse por selección de grupo: una población sexual siempre tendrá multitud de cerraduras, mientras que los miembros de un clon asexual tendrán todos la misma. Así, un parásito que dé con la llave que abre la cerradura del clon asexual puede exterminar en poco tiempo la población entera, mientras que en el caso sexual siempre habrá cerraduras que el parásito no podrá abrir con tanta facilidad.
Las especies sexuales disponen de un banco de cerraduras del que no disponen las especies asexuales. En términos más técnicos, en las especies sexuales hay heterosis y polimorfismo, lo que quiere decir que tanto en el nivel poblacional (polimorfismo) como en el nivel individual (heterosis) hay una variedad de alelos para cada gen. La presión selectiva que mantiene este polimorfismo es, casi con seguridad, las enfermedades infecciosas. La dinámica darwiniana más plausible para la coevolución llave-cerradura es lo que en genética de poblaciones se conoce como selección negativa dependiente de la frecuencia. Tan pronto como una cerradura se hace poco frecuente en la población, la llave correspondiente también disminuye de frecuencia, y en tales condiciones la infrecuencia se convierte en una ventaja selectiva. Esta ventaja pasa continuamente de un alelo a otro (de una cerradura a otra), lo que contribuye a mantener el polimorfismo. Los precedentes de esta idea se remontan al gran genetista inglés J. B. S. Haldane, quien fue el primero en advertir que el parasitismo, más que la predación, podía promover el polimorfismo génico, sobre todo si el éxito incrementado del parásito en infectar una nueva variedad de huésped va aparejado a un éxito reducido contra la variedad antigua, lo que sería congruente con la dinámica de llaves y cerraduras[28].
Pero incluso la recombinación génica derivada del sexo puede ser insuficiente para vencer la inercia evolutiva poblacional en la carrera contra los parásitos cuando el tiempo de generación del huésped es muy largo en comparación con el de sus parásitos. El sistema inmunitario es una innovación reciente a escala evolutiva, inventada por los reptiles y ausente en anfibios, peces e invertebrados. Básicamente consiste en una batería de unos diez millones de tipos de glóbulos blancos, cada uno con una «cerradura» (anticuerpo) correspondiente a una «llave» (antígeno). Si la llave encaja en la cerradura, el glóbulo blanco correspondiente se multiplica frenéticamente para producir una fuerza de choque contra los cuerpos extraños. No hay un gen para cada anticuerpo, sino que cada «cerradura» viene dada por una combinación de proteínas modulares, lo que permite obtener una gama muy amplia de anticuerpos a partir de un único complejo de genes. De este modo el sistema inmunitario puede anticiparse a los antígenos cambiantes de los parásitos.
Para evitar que el sistema inmunitario ataque las células del propio organismo, éstas se reconocen entre sí mediante una contraseña (el llamado «antígeno principal de histocompatibilidad»). Así, para salirse con la suya, el parásito puede adoptar una de las siguientes estrategias: infectar a otro huésped por contagio antes de que el sistema inmunitario de su portador reaccione y lo elimine, o modificar sus propios antígenos a menudo, o esconderse dentro de las células del huésped, o adivinar su contraseña, y aquí es donde vuelve a entrar en juego el sexo. Los genes de histocompatibilidad, responsables tanto de las contraseñas celulares como de la susceptibilidad a las infecciones, son altamente polimórficos (con más de cien alelos cada uno en el caso humano, de los que cada persona posee una combinación única), un polimorfismo cuyo mantenimiento requiere del sexo[29]. Así pues, aunque evolucionara para suplir la insuficiente adaptabilidad darwiniana de los organismos de tiempo de generación largo, el sexo sigue siendo necesario para garantizar la eficiencia del sistema inmunitario.
Sexo y capacidad de anticipación
La respuesta adaptativa de las especies a los cambios de su entorno físico y, sobre todo, biótico requiere adaptabilidad. A una forma de vida cuyo ciclo biológico se completa en cuestión de días, la variabilidad fruto de errores de copia y otras fuentes de mutación aleatoria puede bastarle para proporcionar a sus poblaciones una adaptabilidad suficiente para evitar la extinción, sobre todo si la impredecibilidad del entorno obliga a producir el mayor número posible de descendientes en el menor tiempo posible; en esas condiciones la reproducción asexual será a menudo una estrategia más conveniente que el sexo meiótico biparental. Pero cuando el tiempo de generación se alarga, la inercia evolutiva de las poblaciones aumenta, lo que hace necesaria una fuente de variación suplementaria que evite la pérdida de adaptabilidad darwiniana. La recombinación génica permite obtener genotipos nuevos a partir del acervo génico existente sin tener que esperar a que surjan novedades por mutación, lo que puede explicar la prevalencia de la sexualidad en las especies con tiempos de generación largos. En consonancia con esta idea, un estudio ya clásico de los biólogos Austin Burt y Graham Bell sobre una amplia variedad de mamíferos ha permitido constatar que la magnitud de la recombinación génica (el principal efecto del sexo) por entrecruzamiento (es decir, el intercambio de genes entre cromosomas homólogos durante la meiosis) guarda una correlación nula con el tamaño de la prole, baja con el tamaño corporal y alta con la edad de la madurez sexual (figura 2.1). En otras palabras, la magnitud de la recombinación génica es mayor en las especies de vida larga y desarrollo lento que en las de ciclo vital corto, con independencia del tamaño o la fecundidad[30]. Se comprende así la intrigante correlación entre sexualidad y complejidad organísmica. En realidad, esta correlación es acausal: el parámetro relevante no es la complejidad en sí, sino el tiempo de generación. Lo que ocurre es que las formas de vida más complejas (como las aves y los mamíferos, atados a la reproducción sexual) suelen ser también las que tienen tiempos de generación más largos.
Lo que aporta el sexo a los organismos de tiempo de generación prolongado es una ganancia de independencia de la incertidumbre del entorno. (Para una formalización de este concepto en los términos de la teoría de la información, véase el apéndice matemático). A fin de mantener su identidad con independencia de las veleidades del entorno y evitar la extinción, un sistema vivo no puede limitarse a aislarse del mundo, porque todo sistema aislado tiende al equilibrio termodinámico (es decir, la muerte). Estar vivo implica mantenerse en un estado estacionario alejado del equilibrio termodinámico, lo cual requiere un intercambio de información entre sistema y entorno que garantice la adaptabilidad. Ahora bien, ¿qué ocurre cuando la adaptabilidad del sistema vivo es insuficiente para proporcionarle la capacidad de anticipación que requiere un entorno impredecible? En tal caso el sistema vivo no podrá persistir sin sacrificar su identidad. Deberá instalarse en un nuevo estado estacionario compatible con la nueva estructura del entorno. En otras palabras, para recuperar su independencia de la incertidumbre del entorno deberá adoptar una nueva identidad, aunque ello implique la emergencia de una individualidad de orden superior. (Inversamente, si la incertidumbre del entorno disminuye, entonces el sistema vivo puede permitirse sacrificar parte de su adaptabilidad en aras de la eficiencia. Para circular por el trazado liso, casi recto y predecible de una autopista, un todoterreno es una opción innecesariamente cara y poco eficiente en comparación con un deportivo ligero y aerodinámico, capaz de alcanzar más velocidad con menor gasto de combustible, aunque este último será más sensible a los baches imprevistos. De ahí que la especialización y simplificación de los organismos tienda a aumentar con la predecibilidad del entorno. Pero, cuanto más se especializa, más frágil se vuelve el sistema vivo ante cualquier perturbación inesperada del mundo que lo rodea).
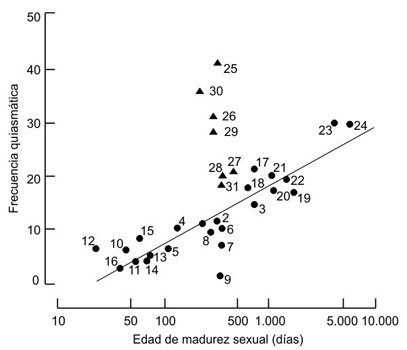
Figura 2.1. Frecuencia de quiasmas (puntos de entrecruzamiento cromosómico, una medida de la recombinación génica meiótica) en función de la edad de madurez sexual (una medida del tiempo de generación) en una amplia muestra de mamíferos. Los círculos corresponden a especies salvajes y los triángulos a especies domesticadas (no incluidas en el cálculo de la recta de regresión). Obsérvese que las especies domesticadas tienen tasas de recombinación aumentadas, lo que seguramente se debe a que han estado sometidas a una selección direccional particularmente intensa. Especies salvajes: 1, Dasyuroides byrnei; 2, Dasyurus viverrinus; 3, Sarcophilus harrisii; 4, Sminthopsis crassicaudata; 5, Perameles gunnii; 6, Isoodon macrourus; 7, Dasypus novemcinctus; 8, Oryctolagus cuniculus; 9, Thomomys bottae; 10, Cricetus cricetus; 11, Mesocricetus auratus; 12, Lagurus lagurus; 13, Meriones unguiculatus; 14, Apodemus sylvaticus; 15, Rattus norvegicus; 16, Mus musculus; 17, Cebuella pygmaea; 18, Saguinus oedipus; 19, Macaca fuscata; 20, Macaca mulatta; 21, Macaca nemestrina; 22, Cynopithecus niger; 23, Pan troglodytes; 24, Homo sapiens. Especies domesticadas: 25, Canis familiaris; 26, Felis catus; 27, Sus scrofa; 28, Bos taurus; 29, Capra hircus; 30, Ovis aries; 31, Equus caballus. (De Burt y Bell, 1987).
¿Cómo puede «inventar» la evolución nuevos estados estacionarios? Hay dos modos fundamentales de innovación evolutiva: mutación y asociación[31]. Cualquier mutación que tenga algún efecto fenotípico implica un eventual nuevo estado estacionario. Las innovaciones así surgidas pueden difundirse después si la selección natural favorece su multiplicación. El otro modo de crear nuevos estados estacionarios es mediante la integración de dos o más estados estacionarios separados en uno solo. En otras palabras, la asociación de individualidades para crear una nueva individualidad a un nivel de organización superior. Las asociaciones viables pueden ser entre individualidades idénticas o emparentadas (sociedades en sentido amplio) o entre individualidades no emparentadas (simbiosis). A diferencia de las mutaciones, las asociaciones permiten ascender en la jerarquía de niveles de organización de la materia viva. Las individualidades de orden inferior (células, organismos) sacrifican parte de su independencia a cambio de pertenecer a una totalidad más independiente de la incertidumbre del entorno que sus integrantes por separado.
La identidad colectiva de una individualidad construida por asociación puede ser tan robusta que su nueva cohesión se haga irreversible. Es el caso de la pluricelularidad y, en el nivel jerárquico inmediatamente superior, las sociedades de insectos. (En el caso de la simbiosis, la interpretación es esencialmente la misma: la identidad conjunta es más independiente de la incertidumbre del entorno que cada simbionte por separado. Aunque la dependencia mutua de los simbiontes puede aumentar, el conjunto es más independiente. Un buen ejemplo es el de los líquenes, asociaciones de un alga y un hongo capaces de sobrevivir en medios muy pobres, porque el alga produce alimento con los nutrientes reciclados por el hongo a partir de sus productos metabólicos de desecho). Los logros más sofisticados de la evolución biológica (como el sistema inmunitario o el aparato cognitivo capaz de aprender de la experiencia y resolver problemas) se comprenden bien a la luz del criterio de independización creciente de la incertidumbre del entorno. Como argumentaré a continuación, la evolución del sexo también admite una interpretación en los mismos términos.
La idea de identidad individual implica un conjunto de propiedades que permiten a la individualidad viva ser idéntica sólo a sí misma. En los organismos asexuales, cuya progenie es un clon, esta identidad viene dada por el genotipo del organismo individual. Pero los organismos sexuales pierden su identidad genotípica al reproducirse. El sexo permite mantener un acervo de alelos amplio, compartible por el conjunto de la población, lo que incrementa la gama de genotipos posibles por recombinación génica y, con ello, la capacidad de anticipación del sistema vivo a los cambios del entorno (en particular el entorno biótico). Pero esta ganancia de capacidad de anticipación implica renunciar al mantenimiento de la identidad genotípica, porque el sexo desbarata los genotipos parentales y los confunde en la descendencia. De ahí que, desde la perspectiva radicalmente individualista del neodarwinismo ultraortodoxo, la reproducción sexual plantee una paradoja evolutiva.
Sexo y unidades de selección
El pensamiento darwinista gira en torno al individuo, sobre el cual se ejerce la selección natural. Son los individuos los que dejan más o menos descendencia según la adecuación de sus rasgos físicos y de comportamiento a sus condiciones y modo de vida. El individuo es la unidad de selección. Los organismos individuales no obran para perpetuar su especie o mantener el equilibrio ecológico, sino para perpetuar su propia estirpe. Toda armonía o estabilidad de orden superior es un producto secundario de este hecho. Pero, a diferencia de los genetistas de poblaciones, los naturalistas y los ecólogos siempre han tendido a pensar más en términos de selección supraindividual. El propio Darwin apeló al bien de la comunidad para explicar la existencia de castas estériles en diversas especies de insectos sociales. A comienzos de la década de 1960, el escocés Vero Wynne-Edwards se atrevió a desafiar abiertamente la ortodoxia darwinista al proclamar que la unidad de selección es el grupo y no el individuo[32]. Su planteamiento era el siguiente: si los individuos sólo atienden a su propia reproducción, ¿por qué hay tantas especies que mantienen sus poblaciones dentro de unos límites ajustados a los recursos disponibles? Para el darwinismo ortodoxo esto tenía una explicación obvia: la limitación de los recursos y las presiones selectivas del clima y la depredación recortaban cualquier exceso de población. Pero esta respuesta no satisfacía a Wynne-Edwards, quien argumentó que los grupos en los que evolucionara alguna suerte de altruismo demográfico conseguirían mantenerse tanto en los buenos como en los malos tiempos, lo cual sería favorecido por la selección natural. Así pues, la evolución sería más una lucha entre grupos que una lucha entre individuos.
Ante la «herejía» del seleccionismo de grupo, los guardianes de la ortodoxia contraatacaron demostrando que los ejemplos de altruismo aportados por Wynne-Edwards o bien eran reinterpretables en términos individualistas o bien eran producto de lo que William Hamilton llamó «selección de parentesco»[33]. En este último caso, los genes del altruista se perpetúan indirectamente a través de los parientes que se reproducen a su costa. Aunque sacrifique su propia reproducción, el altruista obra en su propio interés genético, porque con su conducta contribuye a propagar sus propios genes más de lo que lo haría reproduciéndose por su cuenta. Es lo que los evolucionistas llaman «aptitud inclusiva». Las conductas altruistas y la ayuda mutua serán favorecidas por la selección natural siempre que se propaguen los genes compartidos por los individuos emparentados, aun a costa de la reproducción de los altruistas. En última instancia, pues, la selección de parentesco no es más que una variante de la selección individual darwiniana; o así quisieron verlo los defensores de la ortodoxia.
Pero el argumento de que la selección natural puede favorecer una suerte de reproducción transindividual es un arma de doble filo. El primero en apreciar las implicaciones de esta idea fue el norteamericano George Williams. Las reflexiones de Williams en torno al problema del envejecimiento (que adquiere sentido evolutivo cuando se interpreta como una obsolescencia genéticamente planificada) le llevaron a subrayar que para la selección natural lo que cuenta no es la supervivencia del individuo, sino la de los genes del individuo. Aunque las presiones selectivas se ejercen sobre los organismos, lo que de hecho se selecciona son sus genes, pues sólo éstos pasan a la siguiente generación. Basándose en este argumento, Williams propuso que la auténtica unidad de selección no es el organismo individual, sino el gen[34].
Si la intención de Wynne-Edwards había sido desplazar al individuo de su posición central en el pensamiento darwinista, en cierto modo se salió con la suya, aunque justo al revés de como él había pretendido. El nuevo paradigma insiste en que los organismos obran no en interés propio, sino en interés de sus genes. Esta puntualización puede parecer ociosa, pero no lo es tanto si se piensa que el bien del organismo individual no siempre es compatible con la óptima propagación de sus genes. En caso de conflicto, es el interés genético el que prima. A veces la supervivencia de los genes requiere que los individuos se sacrifiquen por su descendencia o por sus parientes cercanos. Pero todas estas conductas se han seleccionado porque contribuyen a la perpetuación de los genes del altruista de turno.
El principal paladín de este seleccionismo génico es Richard Dawkins, autor de El gen egoísta, quien ha llegado a afirmar que los organismos no son más que «máquinas de supervivencia» para sus genes. Los genes que persisten son precisamente aquéllos que crean máquinas de supervivencia capaces de perpetuarlos[35]. Los genes se hacen así depositarios de las propiedades que antes caracterizaban a los organismos individuales: se multiplican autocopiándose, varían por mutación y compiten entre sí de modo que las variantes más aptas aumentan de número a expensas de las menos aptas. De esta manera el seleccionismo génico ha restaurado el proverbial individualismo del discurso darwinista, aunque, paradójicamente, lo ha hecho a costa de la individualidad organísmica.
También aquí el sexo tiene un papel principal. Si no existiera la reproducción sexual, el seleccionismo génico no supondría ninguna ruptura conceptual con el darwinismo clásico. Los organismos asexuales producen clones de sí mismos, de manera que la identidad genética del individuo (su genotipo) se perpetúa en su descendencia. Aquello sobre lo que se ejerce la selección (el organismo individual) es lo que pasa tal cual a la generación siguiente, de manera que no hay ambigüedad alguna en cuanto a la unidad de selección. Pero los organismos que se reproducen sexualmente pierden sin remedio su identidad al reproducirse. Al recombinarse los genotipos de ambos progenitores, sus identidades genéticas se confunden en la descendencia, de manera que los hijos nunca son genéticamente idénticos a sus padres. Las individualidades de los organismos sexuales son transitorias e irrepetibles; en cambio los genes (es decir, los paquetes de información codificada en el ADN) son potencialmente eternos. La reproducción sexual hace que selección génica y selección individual dejen de ser conceptos equivalentes, lo que, a ojos de los seleccionistas génicos, justifica el cambio de unidad de selección. (No obstante, hay que señalar que, aunque la reproducción sexual no es conservativa en cuanto al genotipo, sí lo es en cuanto al genoma, el conjunto de genes que define la especie. La transitoriedad de los genotipos individuales no impide que nuestros hijos sean seres humanos como nosotros, y no chimpancés o lagartijas. Aunque pensar en términos de selección de genes —o, mejor, de alelos— pueda ser conveniente para abordar algunos problemas evolutivos, el nivel organísmico sigue ocupando una posición central en el pensamiento evolucionista).
A mi juicio, la cuestión de la unidad de selección (como la cuestión del valor adaptativo del sexo) es en buena medida un falso problema. Aunque el individualismo en el nivel de los genes no necesariamente implica individualismo en el de los organismos, los seleccionistas génicos tienden a olvidar este punto, quizá porque la principal motivación de esta corriente de pensamiento ha sido reafirmar el individualismo radical del darwinismo clásico frente al seleccionismo de grupo. Pero lo cierto es que el seleccionismo génico es perfectamente compatible con el seleccionismo de grupo (¿qué otra cosa, si no, está implícita en la noción de selección de parentesco?)[36]. Esta cuestión es especialmente relevante para el problema del sexo. No en vano el propio Ronald Fisher, uno de los padres del neodarwinismo, ya reconoció desde el principio que la evolución de la reproducción sexual requería una interpretación en términos de selección de grupo.
Sexo como altruismo génico
Puede que los parásitos proporcionen la anhelada solución de la paradoja del sexo: la aparente ausencia de una ventaja adaptativa a corto plazo que compense los teóricos beneficios inmediatos de la alternativa asexual. Pero, aun aceptando que la reproducción sexual pueda ser adaptativa incluso a una generación vista, sigue siendo difícil de conciliar con el pretendido «egoísmo» de los genes. Los adeptos a una concepción radicalmente individualista de la selección natural suelen obviar el hecho de que en la naturaleza no existen genes que vayan por libre. Hasta los virus más simples son asociaciones de decenas de genes que cooperan para reproducirse, y no hay ningún organismo de reproducción autónoma cuyo genoma contenga menos de 400 genes (el tamaño genómico de los micoplasmas más pequeños). En el nivel génico, el más fundamental de todos los niveles de selección, la cooperación es la norma y no la excepción.
La omnipresencia de los genes cooperantes en el mundo vivo sugiere que el egoísmo a ultranza no es una estrategia evolutivamente estable ni siquiera al nivel génico. Para salvar los muebles, Richard Dawkins, el padre de la metáfora del gen egoísta, argumenta que la cooperación a cualquier nivel es una resolución particular del conflicto de intereses fundamental entre los genes individuales que se ve favorecida por el hecho de que el entorno de cada gen está dominado por otros genes[37]. En otras palabras, los genes cooperan, sí, pero sólo porque obtienen un beneficio individual de ello; es un estilo de cooperación «anarquista» en el que cada gen atiende sólo a sus propios intereses egoístas.
Si todo lo que quiere decir Dawkins es que los genes nunca obrarán en perjuicio de su propia perpetuación, entonces es difícil estar en desacuerdo con él. Se puede criticar su uso (o abuso) del término «egoísmo». Pero, cuestiones semánticas aparte, es innegable que los genes también compiten entre sí de manera excluyente. En lo que Dawkins se equivoca es en generalizar el principio de la exclusión competitiva (lo que él llama egoísmo genético) más allá de su dominio de aplicación. La competencia al nivel génico es fundamentalmente una competencia entre alelos (es decir, variantes de un mismo gen), no entre genes no homólogos. Editores e impresores cooperan en la producción de libros, y ciertamente lo hacen en interés propio, pero una cosa son las relaciones comerciales entre empresas de distintos gremios y otra muy distinta la competencia entre las editoriales, o entre las imprentas, por dominar el mercado. La competencia fundamental es la que se entabla entre alelos que aspiran al mismo puesto (un mismo locus cromosómico) en la sociedad de los genes. Individualismo y cooperación son principios evolutivos compatibles, porque rigen a niveles distintos.
Así pues, la cooperación al nivel genómico no compromete la concepción individualista de la selección al nivel de los alelos aspirantes a ocupar un mismo locus cromosómico. Pero la reproducción sexual sí que la compromete, y mucho, porque implica que dos alelos de cada gen, uno materno y otro paterno, no necesariamente idénticos, se dan la mano y pasan juntos a la siguiente generación. Al hacerlo así, los genes renuncian al beneficio selectivo inmediato de excluir a sus competidores directos, por lo que el sexo puede contemplarse como una forma de altruismo recíproco al nivel génico.
Uno de los ejemplos favoritos de Dawkins es el alelo t del ratón, un gen que favorece su propia propagación a expensas de los otros alelos de su mismo locus. Tales genes se conocen como distorsionadores de la segregación, porque distorsionan la segregación mendeliana de los caracteres en virtud de lo que se conoce como «impulso meiótico», un proceso que sesga la meiosis para favorecer la representación de un genotipo concreto en la descendencia. El alelo «egoísta» se segrega en los heterocigotos con una frecuencia muy superior a la mitad esperada, lo que le confiere una mayor aptitud darwiniana al nivel de la población de gametos, contrarrestada al nivel genotípico por la letalidad, viabilidad disminuida o esterilidad de los homocigotos para el distorsionador. Esta situación conduce a un polimorfismo génico que puede durar mucho tiempo (más de tres millones de años en el caso de los ratones)[38].
Lo que hacen los distorsionadores de la segregación es menoscabar la heterosis y, con ello, revertir el principal resultado del sexo: la producción de genotipos nuevos por recombinación génica. Pero tales alelos «tramposos» son raros[39]. Si fuera cierto que los genes en general se comportan de manera egoísta en el sentido radical de Dawkins, entonces los distorsionadores de la segregación deberían ser la norma y no la excepción. Si convenimos en que el gen es la unidad fundamental de selección, tal como defiende el propio Dawkins, entonces debe admitirse que, al nivel más fundamental de la materia viva, la cooperación se impone al egoísmo excluyente incluso entre alelos que deberían competir por un mismo locus cromosómico.
Sexo y selección de grupo
Cuando se tiene presente el entorno biótico siempre cambiante de los organismos, la tesis de que el valor selectivo de la sexualidad reside en la generación de variabilidad genotípica vuelve a cobrar fuerza. Lo que ha venido confundiendo a los neodarwinistas ortodoxos es que, desde el punto de vista del progenitor individual, la reproducción sexual no parece tener ningún valor selectivo inequívoco que compense los presuntos beneficios inmediatos de la opción asexual. Pero, como ya sugiriera Weismann, el sexo no incrementa directamente la aptitud media, sino la varianza de la aptitud y, por ende, la respuesta a la selección. En otras palabras, los individuos que se reproducen sexualmente tienen progenies más variables, lo que posibilita que su descendencia se adapte más deprisa. De acuerdo con esta idea, si tenemos dos poblaciones, una sexual y otra asexual, por lo demás idénticas, la población sexual acabará desplazando cualquier clon asexual porque la recombinación génica meiótica incrementa la eficacia de la selección. Ahora bien, lo que se selecciona no es un clon de aptitud óptima, sino un clado (es decir, un conjunto de organismos relacionados por vía de descendencia) con un acervo génico compartido. En otras palabras, los beneficios del sexo se recogen a los niveles de clado y de gen (ambos unidades de selección en el sentido de Williams)[40].
El sexo no reporta ningún beneficio directo al individuo que se reproduce sexualmente. Esto no es un defecto de la teoría, pero ha sido contemplado como tal porque la lógica weismanniana se aparta un tanto de la ortodoxia darwinista, con su énfasis en la individualidad organísmica. De ahí que se la haya criticado por requerir una selección de grupo, y se haya objetado que la evolución de la reproducción sexual debe explicarse sobre la base de alguna ventaja selectiva individual e inmediata. Pero esta objeción es más teórica que empírica. Para empezar, como evidencian los casos del impulso meiótico o del altruismo ligado a la selección de parentesco, los individuos no tienen por qué beneficiarse directamente de las adaptaciones que contribuyen a la proliferación de los genes de los que son portadores. Además, el hecho de pasarse a la reproducción asexual no tiene por qué traducirse en un incremento de la descendencia efectiva sin más. Aparte de que las presiones selectivas no son parámetros fijos, como suelen asumir los modelos matemáticos, en la mayoría de situaciones reales la fecundidad depende sobre todo de la inversión parental femenina y de la disponibilidad de recursos, así que no está claro que una hembra partenogenética pueda dejar mucha más descendencia que sus competidoras. En cualquier caso, la diferencia difícilmente sería lo bastante significativa para que su progenie desplazara a las hembras sexuales en las generaciones sucesivas antes de que alguna crisis (en la forma de una epidemia causada por un virus mutante o cualquier otro patógeno, por ejemplo) volviera a poner las cosas en su sitio. El sexo no necesita ser ventajoso a corto plazo, basta con que no sea demasiado desventajoso.
Por otra parte, el argumento del doble coste del sexo aduce que las poblaciones de hembras partenogenéticas son el doble de fecundas que las poblaciones formadas por machos y hembras. Ahora bien, ¿no habíamos quedado en que la selección natural se ejerce sobre los individuos, no sobre las poblaciones? Apelar a la fecundidad diferencial de las poblaciones es incurrir en un seleccionismo de grupo (precisamente lo que más detestan los darwinistas ultraortodoxos). Pero la selección de grupo (o, para ser más precisos, de clado) es justo lo que hay implicado en la competencia entre especies sexuales y clones obligadamente asexuales. En realidad, los machos no suelen ser un factor limitante de la fecundidad femenina, y cuando lo son es porque aportan inversión parental además de espermatozoides, de manera que prescindir del sexo no reporta a las hembras ninguna ventaja añadida. Es más, como veremos en su momento, una hembra sexual puede servirse del mayor potencial reproductivo masculino para propagar más eficazmente sus propios genes (a través de su progenie masculina), cosa que no puede hacer una hembra partenogenética (que sólo engendra hijas).
Sexo progresista y sexo conservador
Para dar por resuelta la paradoja del sexo, nos falta un último trámite. Como hemos visto, los organismos sexuales pierden su identidad genotípica individual al reproducirse, de manera que la individualidad sobre la que presuntamente se ejerce la selección (el organismo) no se corresponde con la identidad que se perpetúa. Para restaurar la correspondencia entre la individualidad objeto de selección y la identidad que pasa a la generación siguiente hay que definir una individualidad supraorganísmica tal que los genes seleccionados caractericen la identidad colectiva que se perpetúa. En el caso de las sociedades de insectos con castas de obreras estériles, la paradoja aparente se resuelve trasladando la individualidad al enjambre entero, que puede contemplarse como una suerte de organismo difuso. Cuando se adopta esta perspectiva, el que las obreras deleguen su propia reproducción en unos cuantos machos y hembras fértiles no resulta mucho más paradójico que el hecho de que las células de nuestros cuerpos deleguen su propia reproducción en los óvulos y espermatozoides producidos por nuestras gónadas.
¿Cuál es, entonces, la individualidad relevante a efectos selectivos cuando de reproducción sexual se trata? Ya no son el macho o la hembra individuales, sino el grupo mínimo constituido por una pareja de progenitores (de cuyo éxito reproductivo conjunto depende la continuidad genética de ambos). Y la identidad que se perpetúa ya no es la genotípica, sino la definida por un genoma parental común y un acervo de alelos conjunto; en otras palabras, la identidad de especie.
La invención evolutiva del sexo meiótico implica una independencia nueva por asociación, porque los progenitores (al depender unos de otros para reproducirse) sacrifican parte de su independencia individual en aras de una mayor independencia conjunta. Pero, una vez dado este salto evolutivo, el sexo meiótico es en esencia conservador, porque consiste en modificar constantemente el genotipo para mantener invariante la identidad genómica (o, por así decirlo, cambiarlo todo para que nada cambie). En esto el sexo meiótico se diferencia radicalmente del sexo procariótico, que a menudo es «progresista». En los procariotas, cuya sexualidad está disociada de la reproducción, la transmisión de genes es horizontal (entre individuos de la misma generación, y no de padres a hijos) y unilateral (un donante cede parte de su genoma a un receptor). Puesto que implica una cesión de genes y no un intercambio simétrico, cada acto sexual procariota viene a ser un pequeño salto evolutivo, porque el donante pierde una parte de su genoma y el receptor gana genes nuevos, lo que implica que ambos pueden emerger del acto sexual consumado con una nueva identidad genómica[41]. Pero, como ocurre con las mutaciones, la individualidad relevante a efectos selectivos sigue siendo el organismo (en este caso unicelular). Así pues, el sexo procariótico, a diferencia del meiótico, no implica una individualidad supraorganísmica.
El mundo vivo no es una confusión indiferenciada de criaturas. La variedad de formas vivas no es continua, sino que se distribuye en categorías específicas. ¿Por qué hay especies? Ésta es la pregunta que Darwin intentó responder (sin conseguirlo del todo). Las individualidades supraorganísmicas que llamamos especies son intuitivamente diferenciables incluso por el profano. No hace falta ser un naturalista profesional para distinguir de un vistazo un gorrión de un pinzón, o una lagartija de un geco. Pero, en biología, las especies no se definen sólo por una lista de rasgos necesarios para que un espécimen pueda asignarse a una u otra categoría específica. Las especies biológicas se definen, además, como grupos reproductivamente aislados. (Por supuesto, este criterio no es aplicable a las «especies» asexuales). Y es precisamente este aislamiento reproductivo lo que mantiene la identidad de especie.
La pertenencia a una especie no es una cuestión de grado, sino de todo o nada. Las discontinuidades entre categorías específicas son tan básicas para el concepto de especie biológica como los rasgos compartidos. Para los naturalistas predarwinianos como el gran botánico sueco Linneo, introductor de la nomenclatura binomial para clasificar la diversidad de organismos de manera sistemática, el carácter discreto de las especies no representaba un problema. Simplemente eran la obra del Creador, y su identidad se mantenía en virtud de la fidelidad reproductiva. Pero incluso un creacionista como Linneo tuvo que admitir que las especies se transforman, y que pueden llegar a extinguirse. ¿Cómo puede conciliarse la mutabilidad de las especies con las separaciones entre ellas? Si se acepta, como argumentó Darwin, que la evolución es un proceso gradual que procede por pasos imperceptibles, ¿dónde están los eslabones intermedios que deberían conectar todas las especies? Este problema preocupaba sobremanera a Darwin. Podemos imaginar, como hizo él, que los eslabones intermedios existieron en algún momento, pero se han extinguido. El problema es que, cuando los buscamos en el registro fósil, tampoco aparecen. Como es bien sabido, para salir del paso Darwin apeló a «la extrema imperfección del registro geológico».
El origen sexual de las especies
La solución a este rompecabezas quizá resida en el llamado «coste de la rareza». La reproducción sexual tiene un coste intrínseco a bajas densidades de población, debido sobre todo a la disminución del número de parejas potenciales (junto con factores como el incremento de la endogamia, que también pueden tener una contribución significativa)[42]. A medida que las especies se adaptan a su entorno tienden a especializarse en condiciones y parámetros medioambientales restringidos; y a medida que la biota se diversifica, se diferencian nuevas especies, cada una adaptada a una región más limitada del espacio ecológico. En consecuencia, cada vez más especies se agolpan en una región paramétrica reducida. La asunción clave aquí es que cuantas más especies ocupan una región continua del espacio ecológico, más disminuyen sus poblaciones respectivas. Esto es así porque estamos considerando especies muy similares, y el ecosistema sólo puede mantener cierta biomasa de organismos con requerimientos ecológicos parecidos. Así, a medida que las poblaciones de cada especie se enrarecen, el coste de la rareza aumenta, hasta que los beneficios de una adaptación más precisa y eficaz a condiciones cada vez más locales ya no compensan dicho coste. La partición del hábitat cesa, la distribución de especies sexuales se estabiliza y se consuma su segregación. Las separaciones resultantes entre especies son el límite de su similitud, y la consecuencia del coste de la rareza inherente a la reproducción sexual.
Por las razones expuestas, un continuo de especies sexuales, sin discontinuidades perceptibles entre ellas, sería evolutivamente inestable. Para verlo, consideremos una situación de partida con un número incontable de especies, cada una con un nicho ecológico muy reducido y apenas diferenciable de las especies que ocupan nichos ecológicos contiguos, cuyas poblaciones se distribuyen de manera uniforme. Supongamos una fluctuación aleatoria de esta distribución uniforme: la población de una especie cualquiera aumenta al azar y la de otra disminuye en consecuencia, de manera que la biomasa total no cambia. (Podemos asumir que esto no afecta a la aptitud darwiniana si los nichos ecológicos caracterizados por los picos de las curvas de aptitud de ambas especies son contiguos). Puesto que los tamaños poblacionales de partida son ínfimos, el coste de la rareza es exorbitante, de manera que cualquier mínimo aumento del número de parejas potenciales incrementa sobremanera la probabilidad de encontrar una pareja en condiciones. Esto incrementa a su vez la aptitud de la especie cuya población ha aumentado, y a partir de aquí, por un efecto de bola de nieve, las diferencias imperceptibles iniciales se amplifican, hasta que las especies de los nichos contiguos desaparecen. Una especie que inicialmente ocupaba una región minúscula del espacio ecológico ocupa ahora un nicho mucho más amplio, y las especies que ocupaban nichos contiguos se han esfumado, con lo que al final se tienen especies separadas[43].
La delimitación de las «especies» asexuales es un asunto bastante más complicado. Aparte de que no puede aplicárseles el criterio de aislamiento reproductivo, los organismos asexuales no tienen que pagar el coste de la rareza, por lo que es de esperar que sus «especies» proliferen y haya eslabones intermedios por doquier. Las observaciones parecen confirmar esta predicción, ya que la profusión de clones apenas diferenciables es corriente no sólo entre los microbios asexuales, sino entre los organismos pluricelulares que han abandonado el sexo[44]. A modo de ilustración, compárense los casi cien linajes clónicos del gorgojo partenogenético Otiorrhynchus scaber (un coleóptero que infesta los bosques de Suecia y Finlandia) con las seis especies hermanas agrupadas bajo la denominación Anopheles maculipennis (un mosquito que se encuentra por toda Europa)[45]. La diferencia es manifiesta. En el caso sexual, sólo hay seis grupos reproductivos segregados en áreas geográficas amplias, con nichos ecológicos bien diferenciados: dos especies son septentrionales y crían en aguas dulces y salobres respectivamente, otras dos son meridionales y también se diferencian por sus preferencias de salinidad, otra es alpina y la última se restringe a las regiones áridas orientales. En el caso asexual, en cambio, tenemos una profusión de clones entremezclados, con diferencias muy sutiles en sus requerimientos ecológicos.
La proliferación de clones de gorgojos es una situación típica de las formas de vida asexuales: una profusión de ecoespecies, cada una adaptada a los matices más sutiles del entorno, sin más restricciones que la condición trivial de que cada clon debe estar representado por al menos un individuo. En el caso de los mosquitos sexuales, lo que observamos es una segregación de la variedad genérica en un número reducido de unidades básicas (especies) distribuidas en áreas geográficas más amplias y separadas por discontinuidades en el uso de los recursos y el entorno.
El sexo siempre se ha considerado clave para la definición de especie biológica. El intercambio sexual de genes se traduce en un patrón reticulado característico de interconectividad y unidad. La identidad de especie se refiere a la cohesión genómica de los organismos sexuales reproductivamente compatibles. Pero no hay nada en esta unidad genómica que requiera vacíos entre las especies. La distinguibilidad de las especies se refiere a los vacíos en el espacio genómico que a menudo separan una red interconectada (una especie) de otra. Pero, como acabamos de ver, el sexo también tiene un papel clave aquí. Las no linealidades inherentes a la dinámica poblacional de los organismos sexuales son las responsables de la distinguibilidad de las especies. El sexo tiene dos efectos aparentemente opuestos sobre el mundo vivo: unidad y segregación[46].
Las especies sexuales son colecciones de organismos únicos e irrepetibles que existen sólo como depositarios de combinaciones genotípicas transitorias, cada una de las cuales tiene una existencia efímera (a escala evolutiva) desde que nace hasta que es desbaratada por el mismo proceso sexual que la creó. Los organismos nacen y mueren, y entretanto dejan más o menos descendencia en consonancia con la adecuación adaptativa de sus rasgos. Pero, precisamente por causa del sexo, los organismos individuales tienen un papel fugaz en el proceso evolutivo. Los genes, en cambio, son potencialmente eternos. Cambiarlo todo para que nada cambie. De esto va el sexo.