1
¿Por qué existe el sexo?
El ciclo biológico característico de los animales se inicia con la fusión de un espermatozoide y un óvulo en una célula huevo o zigoto, del cual se forma un embrión que se desarrolla hasta convertirse en un organismo pluricelular adulto. Puesto que el óvulo materno y el espermatozoide paterno son portadores de un juego de cromosomas cada uno, el zigoto contiene una dotación génica doble, lo que implica que cada gen estará representado en el nuevo ser por un par de alelos (variantes de un mismo gen) no necesariamente idénticos, uno heredado de la madre y otro del padre. (La excepción son los famosos cromosomas sexuales X e Y, el único par no homólogo; en los mamíferos, un par XX dará lugar a una hembra y un par XY a un macho, mientras que en las aves es al revés). Los biólogos llaman «diploidía» a esta duplicidad cromosómica. Todas las células que nos constituyen son diploides, salvo los óvulos y espermatozoides, que son haploides (lo que significa que tienen un solo juego de cromosomas). En los animales, las plantas, los hongos y otros organismos eucariotas (es decir, constituidos por células nucleadas, a diferencia de los procariotas o bacterias), las fases diploide y haploide se alternan a lo largo del ciclo biológico.
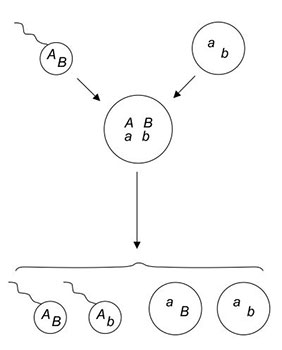
Figura 1.1. Esquema del sexo meiótico. Dos gametos haploides de genotipos AB y ab, procedentes cada uno de un progenitor, se fusionan para dar un zigoto diploide de genotipo Aa/Bb. La meiosis subsiguiente vuelve a producir gametos haploides, recombinantes o no. (De Stearns y Hoekstra, 2000).
La reversión cíclica al estado haploide tiene lugar a través de una meiosis, un modo de división celular que da cuatro gametos (es decir, células sexuales haploides) a partir de una célula diploide (figura 1.1). En los animales la meiosis produce espermatozoides y óvulos destinados a encontrarse y fusionarse para dar huevos fecundados que se desarrollarán en embriones. Las plantas también se desarrollan a partir de embriones diploides. Para completar sus ciclos biológicos generacionales, las células de animales y plantas alternan entre la condición haploide y la diploide. Esta alternancia de las fases haploide y diploide, en forma de meiosis y consiguiente fusión de células sexuales, se da también en los ciclos biológicos de la mayoría de hongos, así como de multitud de algas y protozoos.
Un pasado caníbal
¿Cómo evolucionó el sexo meiótico? Los precedentes evolutivos de esta modalidad reproductiva seguramente se remontan a más de mil millones de años atrás, mucho antes de la aparición de los primeros animales y plantas en el registro fósil. La microbióloga Lynn Margulis, inspirándose en una vieja propuesta de su colega L. R. Cleveland, ha pintado un sugerente cuadro de este proceso evolutivo[1]. Azuzadas por el hambre, las amebas ancestrales que poblaban las charcas anóxicas de la Tierra primigenia se habrían abalanzado unas sobre otras en un holocausto caníbal. Pero en ocasiones la víctima no habría sido digerida del todo por su depredadora, lo que habría dado lugar a quimeras con dos núcleos, que en los casos más afortunados se habrían fusionado, creando células uninucleadas con dos juegos de cromosomas repetidos. En aquellos fangos primigenios, a resguardo del para ellas venenoso oxígeno (un producto de desecho de las recién evolucionadas cianobacterias, responsables de la primera gran crisis medioambiental a escala planetaria), las entidades diploides así formadas habrían resistido la eventual escasez de agua y nutrientes mejor que sus contrapartidas haploides.
Pero tarde o temprano las células diploides habrían tenido que revertir al estado haploide óptimo en condiciones normales. De hecho, en muchas algas y protozoos actuales el sexo meiótico está del todo ausente. Estos organismos simples se reproducen asexualmente por división, sin más complicaciones, y no se molestan en producir un juego de cromosomas superfluo. No hay sexos ni apareamientos. Incluso numerosas algas y hongos facultativamente sexuales, y hasta algunos animales, prefieren reproducirse por vía asexual la mayor parte del tiempo. Sólo se aparean para superar alguna coyuntura difícil, y el producto de la fusión es una estructura resistente, un propágulo cuya misión es experimentar una meiosis para restaurar la fase haploide en cuanto las circunstancias lo permitan. Los ancestros de los organismos pluricelulares animales y vegetales también fueron eucariotas haploides que, en condiciones de tensión medioambiental, se fusionaban por pares formando entidades diploides que revertían al estado simple previo cuando las circunstancias volvían a ser favorables.
Aquellos organismos tenían que resolver un problema: generación tras generación, los pares complementarios debían encontrarse y reconocerse unos a otros para fusionarse y recrear la forma resistente capaz de sobrevivir a la desecación, la congelación o cualquier otra calamidad recurrente. Al principio, las células que se apareaban eran los propios organismos unicelulares. Aún hoy, en muchas algas y protozoos las células sexuales complementarias son indistinguibles. Con el tiempo, y en numerosos linajes por separado, dichas células sexuales fueron diferenciándose en tamaño y forma: de la isogamia se pasó a la anisogamia, un proceso que culminó en la diferenciación entre espermatozoides y óvulos, que a su vez propició la diferenciación de los sexos masculino y femenino. Pero éste es el tema del capítulo 3.
Alternancia de generaciones
Como acabamos de ver, el paso inicial en la evolución de los primeros ciclos biológicos con alternancia de fases haploide y diploide muy bien pudo haber sido el canibalismo. La duplicación del número de cromosomas por fusión sexual requiere una explicación evolutiva, y el canibalismo, frecuente entre los microbios, proporciona un punto de partida. Si la célula devorada, o al menos su material genético, pervive dentro de la devoradora, la asociación resultante constituye un estadio evolutivo intermedio entre la depredación y el sexo. Pero todavía faltan otras piezas del rompecabezas. El canibalismo no consumado ciertamente podía duplicar el número de cromosomas y crear una entidad diploide. Pero esta diploidía debía revertir a la haploidía original. Puede que los quistes y otros propágulos diploides derivados de las fusiones celulares posibilitaran la supervivencia en estado latente, pero, cuando volvían los buenos tiempos, replicar un solo juego de cromosomas era más rápido y barato que replicar dos. Se hacía necesaria una división reduccional, y el mecanismo celular más plausible para su evolución habría sido una demora de la replicación cromosómica en el curso de la división celular normal. De hecho, las asincronías entre la replicación cromosómica y la del resto del cuerpo celular son relativamente frecuentes, y cualquier demora que se tradujera en una reducción del número de cromosomas a la mitad se habría visto favorecida por la selección natural[2].
Aunque la meiosis no deja de ser un acto reproductivo, porque produce cuatro células haploides a partir de un zigoto diploide derivado a su vez de la fusión de dos células sexuales haploides, en los eucariotas simples productores de quistes o esporas resistentes aún no puede hablarse con propiedad de reproducción sexual, porque el zigoto diploide no pasa de ser una fase de transición inactiva que se limita a experimentar una meiosis para restaurar la fase haploide en cuanto las condiciones vuelven a ser favorables. Todos estos organismos se multiplican sin necesidad de sexo en condiciones normales; las fusiones sexuales son sólo un recurso de emergencia. Pero en los ancestros de los organismos pluricelulares complejos la fase diploide fue adquiriendo cada vez más preponderancia a expensas de la fase haploide, hasta culminar en el ciclo biológico de los animales, cuyos zigotos se desarrollan en organismos altamente complejos con tejidos y sistemas de órganos, mientras que los gametos haploides, incapaces de multiplicarse por sí solos, han pasado a ser una mera fase de transición entre generaciones sucesivas de diplontes, de reproducción obligadamente sexual en la mayoría de especies.
Entre ambos extremos evolutivos pueden encontrarse todas las posibilidades intermedias. Así, en el ciclo biológico vegetal típico, la alternancia de fases nucleares va unida a una alternancia de generaciones: un haplonte (el gametófito) produce células sexuales cuya fusión engendra un diplonte (el esporófito) que genera esporas haploides por meiosis, cada una de las cuales germina en un nuevo gametófito. En algunas algas, como Cladophora, el gametófito y el esporófito son morfológicamente indistinguibles y llevan vidas independientes. En los musgos, en cambio, el esporófito es una simple cápsula pedunculada que crece sobre el gametófito (la planta verde con hojitas escamosas) y se abre cuando maduran las esporas que contiene. Este dimorfismo generacional se invierte en los helechos, cuyos esporófitos son plantas vasculares frondosas, mientras que los gametófitos son simples talos que recuerdan una hepática. (Algo parecido ocurre en los hongos, cuyos cuerpos fructíferos diploides productores de esporas —las setas— tienen mayor complejidad estructural que los micelios haploides de los que proceden). La regresión del gametófito es casi total en las plantas productoras de semillas, donde se reduce a unas pocas células dentro del grano de polen (en el caso masculino) o el ovario de la flor (en el caso femenino). Por último, en los animales como nosotros la generación haploide está ausente, y los gametos masculinos y femeninos son producidos por los propios diplontes (sexuados o hermafroditas, según las especies), sin intermediarios haplontes. (Hay que decir que muchos invertebrados alternan la reproducción sexual con la partenogénesis, la gemación u otra vía asexual. Pero esta alternancia de modos reproductivos no implica una alternancia entre una generación haploide y otra diploide)[3].
La ciclicidad es un factor clave en los orígenes de la reproducción sexual. Es más que probable que las primeras formas de vida que adoptaron el sexo meiótico tuvieran que alternar entre una estación favorable a la proliferación y una estación desfavorable (gélida o seca) que pasaban en estado latente. Los embriones pluricelulares diploides de plantas y animales comienzan allí donde acaban los quistes microbianos. Su evolución abrió todo un abanico de posibilidades ecológicas nuevas. La existencia misma de los ancestros de animales, plantas y hongos habría dependido cada vez más de la fusión sexual recurrente; y esta sexualidad muy bien puede ser la secuela de un canibalismo inducido por el hambre y la sed.
¿Por qué hay tanto sexo?
La prevalencia de la reproducción sexual entre las formas de vida pluricelulares sigue turbando a los evolucionistas porque su valor adaptativo no resulta evidente. De hecho, al menos a primera vista, el sexo parece un mal negocio, ya que comporta inversiones y costes considerables con rendimientos inferiores a los de la clonación asexual. Para empezar, está el tiempo y la energía invertidos en encontrar un socio reproductor fiable. Luego está el coste genético, porque cada progenitor lega a cada descendiente sólo la mitad de sus genes. Además, los genotipos parentales se confunden en la descendencia, con la consiguiente pérdida de combinaciones génicas óptimas. Por último, está la reducción del potencial reproductivo. Si una madre partenogenética produce el mismo número de hijos que una madre sexuada y las progenies de ambas son igualmente aptas por término medio, entonces la primera tendrá una aptitud doble que la segunda, porque las hembras sexuales dilapidan la mitad de su potencial reproductivo en producir machos incapaces de poner huevos o gestar crías, de manera que la producción de un número dado de descendientes requiere el doble de efectivos que en el caso asexual. Este coste doble del sexo debería hacer que los individuos sexuados se vieran rápidamente suplantados por los asexuados. La ventaja de las hembras partenogenéticas sólo se minimizaría si los machos contribuyeran de alguna manera a incrementar el potencial reproductivo de sus parejas (compartiendo los costes de la crianza, por ejemplo; pero en la naturaleza los padres que ayudan en la crianza son la excepción y no la regla, así que la contribución paterna no puede explicar la prevalencia del sexo)[4].

Figura 1.2. El doble coste del sexo. En teoría, una población de hembras partenogenéticas (b) produciría el doble de descendientes por generación que una población de machos y hembras sexuales (a), suponiendo que todas las hembras sean igualmente fecundas y su descendencia tenga la misma aptitud darwiniana. (De Fontdevila y Moya, 2003).
A pesar de estos costes, la reproducción sexual es una realidad biológica omnipresente en el mundo eucariota. Hay que decir, no obstante, que lo más habitual es la sexualidad facultativa, sobre todo en los organismos microscópicos o muy pequeños, de tiempo de generación breve y poblaciones que se cuentan por millones de individuos. Muchos de estos organismos se reproducen asexualmente en condiciones normales, y sólo se aparean en coyunturas especiales. La reproducción sexual sólo es obligada en los organismos grandes, de tiempo de generación largo y tamaños poblacionales comparativamente pequeños, como es el caso de los mamíferos, las aves y la mayoría de vertebrados.
El sexo es, en esencia, cruzamiento más recombinación. Cada vástago engendrado sexualmente hereda una combinación de los genes de sus cuatro abuelos repartida en una dotación doble, un juego de cromosomas paterno y otro materno. Cualquiera que sea la ventaja del sexo, cabe pensar que debe de estar ligada a esta mezcla de genes. La hipótesis clásica es que la reproducción sexual se ha seleccionado porque genera variación, lo cual incrementa la adaptabilidad a los cambios del entorno. En las especies sexuales, la progenie de una pareja de progenitores puede compartir mutaciones surgidas en linajes distintos, cosa que no ocurre en las especies asexuales. Una población sexual no tiene que esperar a que se produzcan dos eventos de mutación, cada uno infrecuente de por sí, para que dos mutaciones surgidas por separado confluyan en el genotipo de un mismo individuo. El sexo acelera el proceso evolutivo, y en ello residiría su principal ventaja[5].
El problema con esta explicación clásica es que invoca implícitamente un seleccionismo de grupo, porque demanda que los progenitores compartan de buena gana sus genes a fin de generar la innovación requerida para garantizar la adaptabilidad de la población, renunciando así de manera altruista a perpetuar sus identidades genéticas individuales. Por eso incomoda sobremanera a los neodarwinistas «ultraortodoxos», que la encuentran inconciliable con el teórico individualismo «egoísta» de los actores del drama evolutivo. Si el sexo es adaptativo, objetan, es porque debe ser ventajoso para los individuos, sobre los cuales se ejerce la selección natural darwiniana, no para las poblaciones. Pero entonces, ¿cuál es la ventaja del sexo en términos de éxito reproductivo diferencial? La respuesta definitiva a esta pregunta se ha demostrado tan esquiva que el propio George Williams, el gran patriarca de esta nueva ultraortodoxia, acabó acariciando la idea de que el sexo quizá no sea más que un «accidente congelado» evolutivo[6]. En otras palabras, aunque pasarse a la reproducción asexual pudiera reportar alguna ventaja a efectos de selección individual, en la práctica esto no es evolutivamente factible. Así como los cetáceos siguen respirando aire a pesar de haberse adaptado a vivir siempre en el agua, aun a riesgo de ahogarse, porque es la única manera que tienen de oxigenar su sangre, los organismos pluricelulares están atados a la reproducción sexual porque es la única manera que tienen de producir embriones[7].
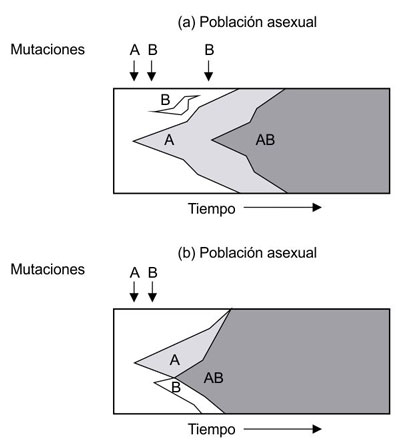
Figura 1.3. Fijación de una combinación ventajosa de mutaciones en organismos con reproducción asexual (a) y sexual (b). En el caso asexual, la mutación B debe darse en organismos previamente portadores de la mutación A, mientras que en el caso sexual las mutaciones A y B pueden surgir de manera independiente y luego confluir en un mismo genotipo AB por recombinación a partir de progenitores portadores de A y de B por separado. (Modelo de Crow y Kimura, en Fontdevila y Moya, 2003).
Pero lo cierto es que la partenogénesis (es decir, el desarrollo espontáneo de un embrión a partir de un óvulo sin necesidad de fecundación previa por un espermatozoide) es una opción reproductiva bastante frecuente en el reino animal, al menos entre los invertebrados. Incluso entre los vertebrados se conocen linajes de hembras partenogenéticas en una veintena de peces, otros tantos anfibios y una treintena de reptiles (casi todos lagartijas y gecos, junto con algún que otro ofidio)[8]. Recientemente se han documentado dos casos de maternidad sin intervención masculina en hembras de dragón de Komodo (un saurio gigante que supera los tres metros de largo y los cien kilos de peso) mantenidas en cautividad[9]. La existencia de reptiles partenogenéticos evidencia que la reproducción asexual es una vía evolutiva accesible también para las formas de vida altamente complejas. Así pues, decir que el sexo es una ligadura histórica insalvable es eludir la cuestión. Pero no faltan propuestas de solución a este dilema.
Sexo reparador
La información genética codificada en el ADN consiste en una doble secuencia de nucleótidos (las «letras» del código genético), cada una complementaria de la otra. La radiación ultravioleta y los radicales libres deterioran el material genético; por eso hay enzimas encargados de su mantenimiento que recorren la doble hélice y reparan cualquier desperfecto, valiéndose de la secuencia complementaria como plantilla para restaurar el tramo dañado. Pero a veces el deterioro local afecta a ambas secuencias (como ocurre cuando las dos cadenas de la doble hélice se fusionan químicamente, lo que impide la transcripción y replicación de la región afectada) y en tal caso los enzimas reparadores tienen que recurrir a una plantilla externa. El sexo proporciona esta copia de seguridad en la forma de una secuencia de nucleótidos intacta procedente de un congénere.
Cuesta explicar por qué las células precursoras de los gametos duplican sus cromosomas y los entrecruzan antes de la reducción meiótica que produce cuatro gametos haploides, si no es para restaurar y «rejuvenecer» la dotación génica destinada a la futura descendencia. Por eso algunos autores consideran que la función primaria de la sexualidad no es otra que la reparación del ADN, y ésta sería la razón de que los organismos sigan apegados a ella a pesar de sus costes[10]. Esto parece poco discutible en lo que concierne a la sexualidad procariota. En la conjugación bacteriana, lo más parecido a la fusión sexual eucariota, una bacteria donante transfiere parte de su genoma a otra bacteria receptora a través de un tubo formado ex profeso. Después el ADN del donante se recombina con el del receptor en los tramos homólogos, y una vez efectuadas las reparaciones pertinentes el material genético sobrante es digerido[11].
Los orígenes de la recombinación génica muy probablemente se remontan a las primeras bacterias, o incluso a las protocélulas de las que se derivaron, y es casi seguro que su función primordial era la reparación del material genético. En un mundo bañado de radiación ultravioleta, sin la capa de ozono que hoy nos protege, aquellas primeras células necesitaban incorporar secuencias de ADN intactas que sirvieran de plantilla para la restauración de las secuencias deterioradas complementarias, y aprendieron a hacerlo a través de la conjugación, la transformación (incorporación de ADN ajeno procedente de congéneres disgregados) y la transducción (intercambio genético mediado por virus). Las células eucariotas, entidades más complejas derivadas de la asociación de procariotas simbiontes, presumiblemente también haploides en su origen, tuvieron que reinventar el sexo para resolver el mismo problema. La solución adoptada fue el sexo meiótico, la fusión de células haploides (singamia) para formar un zigoto diploide, y la subsiguiente meiosis para restaurar la fase haploide una vez efectuada la recombinación génica reparadora.
Como hemos visto, al principio la diploidía era una fase transitoria inactiva entre dos generaciones haploides. Pero la fase diploide fue haciéndose cada vez más preponderante en el ciclo biológico de los organismos pluricelulares a medida que éstos ganaban complejidad. Nuestros cuerpos están formados por células diploides (salvo los óvulos y espermatozoides producidos por nuestras gónadas) y lo mismo vale para casi todos los organismos con los que estamos familiarizados. La razón de la promoción evolutiva de la fase diploide en detrimento de la haploide seguramente tiene que ver con la necesidad de remediar los errores durante la replicación del ADN[12]. Cuanto más complejo es un organismo, más información genética requieren su construcción y funcionamiento. A más genes más ADN, lo que implica más errores de copia y más mutaciones, que casi siempre son perjudiciales.
Una manera obvia de minimizar la tasa de mutación es aumentar la fidelidad de la replicación. Un virus típico como el de la gripe, con menos de diez genes codificados en unos diez mil pares de bases, que cometa un error por cada diez millones de nucleótidos copiados generará una réplica defectuosa de sí mismo por cada mil réplicas exactas. Para conseguir una fidelidad de replicación comparable, una bacteria típica, con unos 4000 genes codificados en alrededor de cinco millones de pares de bases, no puede permitirse más de un error por cada cinco mil millones de nucleótidos copiados. Los eucariotas unicelulares haploides todavía pueden conseguir una tasa de mutación aceptablemente baja a base de minimizar los errores de copia, pero el coste de esta estrategia se hace prohibitivo por debajo de un error por cada cien mil millones de nucleótidos copiados. Esta fidelidad puede bastarle a un protista con menos de 10 000 genes, pero es insuficiente para los organismos pluricelulares complejos, que además de tener que replicar más genes, lo que implica secuencias genómicas más largas, deben repetir el proceso muchas más veces por generación, porque cada nuevo individuo está constituido por millones de células. Por poner un ejemplo, la tasa de mutación para la conocida mosca del vinagre (Drosophila melanogaster) se incrementa en tres órdenes de magnitud, lo que supone una mutación por descendiente. Puesto que mejorar la fidelidad de replicación se hizo evolutivamente inviable, los eucariotas pluricelulares encontraron una solución más simple: mantenerse en estado diploide la mayor parte del tiempo. De esta manera cada gen está duplicado, así que cualquier error fatal en una de las copias (o alelos) pasará inadvertido si la otra copia es correcta. La fase diploide se hizo dominante porque la diploidía enmascara los alelos mutantes disfuncionales e impide que se expresen[13].
Madres vírgenes lesbianas
La diploidía (una consecuencia de la fusión sexual de células haploides) permite disponer de una copia de seguridad del propio genotipo sin tener que aparearse con ningún congénere para obtenerla, ya que todo organismo diploide posee dos copias de cada gen. A efectos de restauración del material genético, por lo tanto, los organismos diploides podrían prescindir del sexo que los creó. De hecho, si de lo que se trata es de recuperar la información genética perdida, parece que la mejor solución es recurrir al propio genoma y no a uno extraño. Así lo hacen numerosas plantas hermafroditas que han renunciado al sexo biparental y han optado por la autofecundación, que viene a ser un apareamiento consigo mismo. La partenogénesis meiótica (el desarrollo de un embrión a partir de un zigoto derivado de la fusión de dos células o núcleos haploides del mismo individuo) documentada en algunos invertebrados y unos pocos vertebrados puede verse también como una autofecundación. Algo distinta es la variante descrita en las lagartijas cola de látigo (Cnemidophorus uniparens), habitantes de los desiertos centroamericanos. Aquí no hay fusión de núcleos haploides, sino que los oocitos (las células precursoras de los óvulos) experimentan una duplicación cromosómica (endomitosis) previa a la meiosis, con lo que al final se obtienen huevos diploides que se desarrollan espontáneamente sin necesidad de autofecundación[14].
El comportamiento de estas lagartijas ha dado mucho que hablar, y es que, aunque la especie es unisexual, lo que quiere decir que sus poblaciones están formadas exclusivamente por hembras partenogenéticas que procrean sin necesidad de machos, no por ello dejan de entregarse al sexo. Las hembras se «montan» unas a otras al modo heterosexual: la participante «activa» se sube al dorso de la «pasiva», le muerde el cuello y se enrosca de manera que los genitales de ambas entren en contacto, permaneciendo así unidas de uno a cinco minutos[15]. El sentido de esta conducta ha suscitado bastante controversia. Para unos, la seudocópula homosexual tiene la función de activar la partenogénesis, mientras que para otros no es más que un epifenómeno, una reminiscencia de la ascendencia dioica de la especie (se sabe que C. uniparens es producto de la hibridación de C. inornatus y C. burti, dos especies dioicas estrechamente emparentadas). Ciertamente, parece un absurdo evolutivo prescindir del sexo reproductor para seguir dependiendo de un simulacro de apareamiento con un congénere (aunque en la naturaleza se ven cosas más raras). En cualquier caso, las lagartijas mantenidas en aislamiento, sin posibilidad de «copular», también ponen huevos partenogenéticos, si bien son menos fecundas[16].
Algunos partidarios de la tesis de que la reproducción sexual es una ligadura histórica ineludible sin sentido adaptativo insisten en que la autofecundación y la partenogénesis meiótica (que no deja de ser una autofecundación) no deben contemplarse como un abandono del sexo, pues se mantiene lo esencial: la meiosis y la singamia. Más que una forma de reproducción asexual, la autofecundación sería «sexo uniparental»[17]. Ahora bien, si nos atenemos a la definición de sexualidad aceptada por los biólogos (es decir, cualquier mezcla de genes procedentes de al menos dos fuentes), entonces la biparentalidad es una condición sine qua non para que pueda hablarse de reproducción sexual. No obstante, hay un aspecto crucial de este «sexo uniparental» que lo acerca al biparental, y es que la progenie producida por autofecundación no es un clon de la progenitora, porque hay recombinación génica (si el genotipo parental es heterocigótico en algún locus cromosómico). En este sentido, la autofecundación se parece más a la reproducción sexual que a la clonación asexual. Pero éste no es el caso de las lagartijas partenogenéticas, porque, aunque mantienen la meiosis, no hay autofecundación, y la progenie así generada sí es un clon de la madre[18]. Por eso considero que, a efectos reproductivos, estos reptiles son claramente asexuales (aunque exhiban una homosexualidad social que parece estimular su fecundidad).
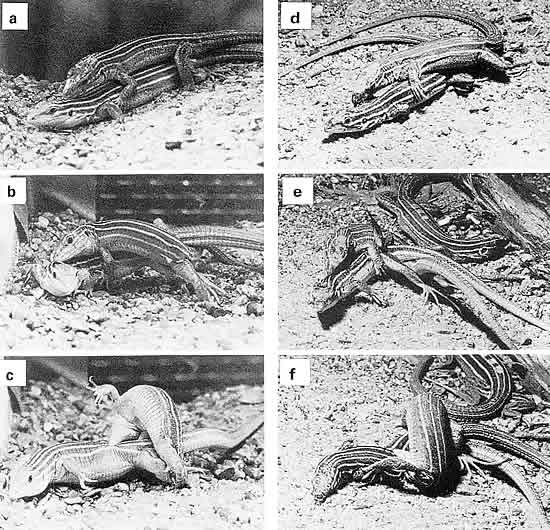
Figura 1.4. El apareamiento de las lagartijas cola de látigo sexuales (en este caso Cnemidophorus inornatus) comienza cuando el macho aborda a la hembra, que si está sexualmente receptiva le deja subirse a su dorso (a). Luego el macho pellizca con las mandíbulas la piel del cuello de la hembra, y maniobra para situar su cola bajo el cuerpo de ésta, a la vez que gira la cadera para oponer su abertura cloacal a la de ella (b). Cuando lo consigue, el macho inserta uno de sus dos hemipenes (todos los lagartos y serpientes tienen penes duplicados) en la cloaca de la hembra, y se contorsiona para trasladar su pellizco del cuello a la región pélvica (c). Esta postura se mantiene de cinco a diez minutos, hasta que el macho suelta su mordisco y se va. Las lagartijas asexuales del mismo género (C. uniparens) exhiben la misma secuencia conductual (d-f). Pero aquí no hay machos (puesto que, desde el punto de vista fenotípico, las lagartijas partenogenéticas son todas hembras), así que es una hembra la que adopta el rol masculino (salvo, obviamente, en lo que respecta a la penetración y la eyaculación). (De Crews, 1994).
Las lagartijas partenogenéticas prescinden de la biparentalidad y de la singamia, pero no de la meiosis, que parece seguir siendo imprescindible para el rejuvenecimiento del material genético legado a la descendencia. De esta manera consiguen restaurar su información genética sin pagar los costes del sexo. En cualquier caso, sólo los organismos unicelulares haploides están obligados a incorporar genes ajenos como plantilla para restaurar los genes propios. En teoría, y aunque no puedan prescindir de la meiosis reparadora, las hembras diploides podrían pasar sin machos, tal como hacen las lagartijas en cuestión.
En resumen, aunque es innegable que la recombinación génica permite restaurar el genoma, y parece claro que los mecanismos moleculares del sexo meiótico se derivan de los mecanismos de reparación del ADN, es más que dudoso que ésta sea la razón principal de la prevalencia de la reproducción sexual en los eucariotas pluricelulares.
El trinquete de Muller
A diferencia del deterioro provocado por la radiación y los radicales libres, los errores de copia durante la replicación del ADN no son reconocidos por los enzimas reparadores. La mayoría de estas mutaciones no tiene efectos fenotípicos significativos, y casi todas las que sí los tienen son más o menos perjudiciales. Pero de tarde en tarde surge una mutación que contribuye de alguna manera al éxito reproductivo de su portador. El sexo no sólo propaga estas mutaciones beneficiosas por la población, sino que permite combinarlas de manera sinérgica, y durante mucho tiempo se pensó que ésa era su principal ventaja.
El problema es que el sexo deshace las combinaciones óptimas de genes tan pronto como las crea[19]. Por eso los biólogos teóricos han vuelto la mirada hacia las mutaciones deletéreas, y han desempolvado un argumento formulado por el genetista Hermann Muller en los años sesenta del siglo pasado, conocido informalmente como «el trinquete de Muller». La idea es que en los linajes asexuales los defectos genéticos se acumulan de manera irreversible. Si fotocopiamos un documento, luego fotocopiamos la copia, y así sucesivamente, la fidelidad de la imagen se irá empobreciendo hasta que el original resulte irreconocible. Si el original se pierde, nunca podremos recuperarlo tal cual era. En la reproducción asexual no hay vuelta atrás. Pero el sexo desbloquea el trinquete y, en virtud de la recombinación génica, permite recuperar genotipos prístinos[20].
Una variante de esta idea pone el énfasis en la necesidad de frenar la acumulación de mutaciones deletéreas, sobre todo en los genomas diploides. Como hemos visto, la diploidía enmascara las mutaciones recesivas, con lo que éstas eluden el escrutinio de la selección natural y se van acumulando de generación en generación, hasta que los genomas se llenan de alelos mutantes recesivos. Dado que estos genes defectuosos no sirven como copia de seguridad para la restauración de la información genética correcta, es conveniente librarse de ellos. En una especie asexual, cada vez que un individuo muere por ser portador de una mutación deletérea la población se libra de esa mutación y sólo ésa. En una población sexual, en cambio, unos individuos heredan muchas mutaciones y otros pocas. Si los primeros mueren, entonces el sexo permite purgar mutaciones del acervo génico de la población, lo que contribuye a mantener desbloqueado el trinquete. Puesto que casi todas las mutaciones son contraproducentes, ahí residiría la ventaja del sexo[21].
Ahora bien, si purgar mutaciones fuera la función principal del sexo, entonces la mejor estrategia sería la autofecundación, porque el apareamiento con uno mismo (la máxima expresión del incesto) es la manera más rápida y barata (porque se evitan los costes de la biparentalidad) de desprenderse de mutaciones recesivas. Para verlo, supongamos que el genotipo parental para cierto locus cromosómico es Aa, donde A es el alelo normal y a un alelo deletéreo recesivo. Entonces (suponiendo una herencia mendeliana simple) el resultado de la autofecundación será:

Es decir, sólo la mitad de la primera generación filial hereda el genotipo heterocigótico parental, mientras que una cuarta parte (los homocigotos AA) se libra del alelo mutante y otra cuarta parte (los homocigotos aa) expresa el gen deletéreo y es eliminada por la selección natural. Es fácil ver que, si se reitera el proceso, los heterocigotos acabarán siendo reemplazados por homocigotos no portadores del alelo mutante.
Pero la autofecundación es más la excepción que la regla entre los organismos que recombinan sus genes por vía meiótica, lo que nos obliga a interrogarnos sobre las ventajas de la fecundación cruzada, o sobre las desventajas de la autofecundación. Para empezar, es bien sabido que la endogamia (de la que la autofecundación es un caso extremo) tiene un coste en términos de éxito reproductivo, porque la descendencia tiende a ser homocigótica, lo que implica la expresión de mutaciones recesivas penalizadas por la selección natural. Al coste de la endogamia debe sumársele lo que se conoce como superioridad heterocigótica. En algunos casos, los heterocigotos tienen fenotipos más adaptativos que los homocigotos, por razones que van más allá del enmascaramiento de mutaciones recesivas. Imaginemos, por poner un ejemplo, que el color de la piel depende de un único gen con dos alelos M y m, tales que los homocigotos dominantes MM fueran melánicos, los homocigotos recesivos mm fueran albinos y los heterocigotos Mm tuvieran un color de piel intermedio, ni negro ni blanco. En tal caso la selección natural penalizaría a los albinos porque serían demasiado proclives al cáncer de piel, pero también a los melánicos porque su epidermis oscura no dejaría pasar luz suficiente para la síntesis dérmica de vitamina D. El fenotipo más apto sería el heterocigótico, que conciliaría de manera óptima la protección del cáncer de piel con la capacidad de sintetizar vitamina D. Éste es un ejemplo imaginario, porque el color de la piel depende de numerosos genes y no de uno solo, lo que permite una gradación fenotípica prácticamente continua desde la palidez total hasta el negro azabache. Pero no faltan ejemplos reales. Uno de los más citados es el caso de la falcemia o anemia falciforme, un síndrome congénito frecuente en las zonas tropicales donde la malaria es endémica, asociado a una mutación recesiva que deforma los glóbulos rojos de la sangre. Los homocigotos recesivos para el gen responsable tienen graves problemas circulatorios y suelen morir antes de la pubertad, mientras que los heterocigotos sólo sufren molestias menores que no les impiden llevar una vida normal. Por otra parte, en virtud de algún efecto fenotípico colateral, los portadores del alelo falciforme son resistentes a la malaria, lo que los hace más aptos que los homocigotos no portadores en un entorno donde la malaria es una causa principal de muerte.
El coste de la endogamia y la superioridad heterocigótica podrían hacer que la selección natural favorezca la fecundación cruzada. Pero en cualquier caso, y aunque la tasa de mutaciones deletéreas sea lo bastante alta en la mayoría de especies sexuales para que la purga mendeliana tenga alguna relevancia, ello sólo demostraría que el sexo es útil para purgar mutaciones, no demostraría por qué se mantiene frente a la alternativa asexual. A ojos de los neodarwinistas ultraortodoxos, las hipótesis basadas en el trinquete de Muller tienen el mismo defecto que la idea de que el sexo acelera la evolución: sus beneficios sólo se recogerían a largo plazo, por lo que la reproducción asexual parece seguir siendo la mejor opción a una generación vista.
La lotería del sexo
Ninguna teoría formulada sólo en los términos de la genética de poblaciones puede explicar de manera plenamente satisfactoria la prevalencia de la reproducción sexual entre los organismos pluricelulares. Por esa razón otros biólogos han propuesto hipótesis de signo más ecológico. La consecuencia más inmediata del sexo es que los hijos no son idénticos a sus progenitores, y éste sería el propósito del sexo según la convención científica imperante hasta los años sesenta. Pero George Williams objetó que esta explicación contraviene la ortodoxia neodarwinista, porque requiere que los individuos renuncien a un beneficio genético inmediato en aras de una ventaja a más largo plazo, algo que, en teoría, sólo podría evolucionar en circunstancias muy particulares. Williams advirtió que, en las especies que alternan ambas formas de reproducción, el sexo suele ir ligado a la dispersión. Muchas gramíneas se reproducen localmente por estolones, pero se dispersan mediante semillas arrastradas por el viento, mientras que los áfidos sexuados desarrollan alas, a diferencia de las hembras partenogenéticas ápteras. La implicación más inmediata es que, si la descendencia está destinada a dispersarse, entonces es mejor que sea variada, porque así es más probable que pueda prosperar en un hábitat impredeciblemente distinto del natal. Otra manera de verlo es que, si la mayor parte de la descendencia que se dispersa está destinada a morir, entonces lo que cuenta no es su aptitud media, sino la probabilidad de que una fracción de la progenie, aunque sea pequeña, esté excepcionalmente bien dotada para salir adelante[22].
El juego de la vida es como una lotería: los organismos asexuales lo apuestan todo a un número, mientras que la reproducción sexual permite diversificar la apuesta, lo que incrementa las posibilidades de acertar. Así pues, el sexo sería la mejor opción reproductiva incluso a corto plazo cuando la progenie debe afrontar un destino incierto. Mientras las condiciones sean favorables y predecibles, es mejor ahorrarse los costes del sexo. Pero cuando la próxima generación debe emigrar o el entorno cambia de manera impredecible, entonces es más conveniente diversificar la descendencia.
Sin embargo, los modelos matemáticos sugieren que el argumento de la lotería sólo tiene peso si el premio es cuantioso. Sólo si una fracción muy pequeña de la descendencia dispersada sobrevive y prospera de manera espectacular vale la pena recurrir al sexo[23]. Pero esta limitación no debilitó la fe de los biólogos en la correlación entre sexualidad e incertidumbre ecológica, hasta que alguien se decidió a verificarla. Los modelos inspirados en la idea de la lotería predicen que el sexo debería imperar allí donde las fluctuaciones del entorno tengan un mayor impacto sobre los organismos: en las latitudes altas más que en las bajas, en las aguas dulces más que en el mar, en los ecosistemas perturbados y jóvenes más que en los maduros, y en las escalas espaciotemporales pequeñas más que en las grandes. Pero esta predicción se ha demostrado demasiado simplista. Los estudios biogeográficos evidencian que la correlación observada entre sexualidad e incertidumbre medioambiental es, al revés de lo esperado, negativa: son los organismos asexuales los que tienden a ser pequeños, de vida corta y residentes en ambientes inestables, mientras que la reproducción sexual obligada es la norma entre las criaturas de ciclo vital prolongado y en los ecosistemas maduros y estables[24].
Es más, los áfidos, las pulgas de agua y otros organismos facultativamente sexuales abandonan la partenogénesis y se pasan al sexo biparental en respuesta no tanto a las fluctuaciones del medio ambiente físico como a la superpoblación. Este hecho ha inspirado otra hipótesis que pone el énfasis en la competencia más que en la incertidumbre del entorno físico. Puesto que los competidores más inmediatos de la mayoría de individuos son sus propios hermanos, cuanto más se diferencien entre sí menos competirán por los recursos locales, y un número mayor de ellos podrá mantenerse. Así como en una economía saturada es conveniente diversificarse, en una situación de competencia intensa por unos recursos limitados es mejor que los hijos no sean clones exactos de los padres, para así atenuar la competencia por el mismo nicho ecológico[25].
El principal inconveniente de esta idea es que, una vez más, huele a selección de grupo. Si cierto genotipo se ha demostrado el más competitivo en la generación previa, en virtud de la superioridad adaptativa del fenotipo correspondiente, ¿por qué no legarlo tal cual a la descendencia? En otras palabras, si algo funciona más que bien, ¿por qué cambiar de modelo? Un reparo más empírico es que la hipótesis de la evitación de la competencia predice que el sexo debería predominar entre los organismos que dejan una descendencia muy numerosa que luego compite entre sí (especies R en la jerga ecológica) más que entre los organismos que controlan su natalidad ajustándola a los recursos disponibles (especies K). Pero no parece que exista ninguna correlación estadística significativa entre estrategias ecológicas y sexualidad[26].