Reexaminaremos ahora, desde un punto de vista biológico, las características de los individuos que componen una raza.
El carácter de una raza es determinado en primer lugar por la herencia. En lenguaje corriente entendemos por herencia el hecho de que el vástago repita la forma de los progenitores sin cambio material, que las características de una serie de generaciones sigan siendo siempre las mismas. Evidentemente esto no es muy exacto, puesto que los descendientes de la misma pareja no serán idénticos en forma ni podrán ser comparados con sus padres ni entre ellos mismos. Cuando consideramos un grupo racial como un todo, suponemos que, a menos que cambien las condiciones, la herencia hará que en generaciones sucesivas se encuentre la misma distribución de frecuencia de formas, o para usar los términos que hemos analizado anteriormente, que la clase, su variabilidad y su término medio permanecerán constantes. Este pensamiento se halla presente en nuestro espíritu al discutir la distribución de tipos raciales. Suponemos que éstos son constantes y continúan inalterables generación tras generación, a no ser que perturbaciones en la población, o quizá, modificaciones de las condiciones exteriores provoquen cambios en la forma corporal.
Podríamos hablar en este sentido le «herencia racial» cuando los rasgos raciales son tan pronunciados que caracterizan a todos los miembros de la raza. Para usar el mismo ejemplo que empleamos antes puede decirse que el cabello rubio, el cutis blanco, los ojos azules o claros, son características hereditarias de la raza sueca, y el cabello negro y rizado, la piel morena y los ojos oscuros, del africano. El hijo de una pareja de suecos no será nunca un africano. Empero, si se compara a los suecos con los alemanes del norte o aún con italianos, se encuentran casos más o menos numerosos en que los rasgos raciales se sobreponen de modo que no es posible hacer una distinción clara. El hijo de una pareja de suecos puede parecerse a un alemán del norte o hasta quizás a un italiano. La expresión «herencia racial» no es aplicable ya, en cuanto significa una determinación de la forma corporal de todos los individuos de la raza.
Hay muchos rasgos que en razas remotas son tan semejantes que tal superposición ocurre con frecuencia. El tamaño del cerebro, la estatura, el peso y el tamaño y forma de varios órganos internos son de este carácter, de manera que respecto a ninguno de ellos podemos referimos como a la herencia racial, como determinación de rasgos que permitan reconocer al individuo como miembro de la raza. Es de capital importancia subrayar que cuando tal superposición existe, una persona de una forma dada que pertenece a una población no es biológicamente, o mejor dicho, genéticamente idéntica a otra persona que tiene las mismas características pero que pertenece a otra población.
Por ejemplo: del estudio de la forma corporal de un número considerable de familias deducimos que las parejas sicilianas cuyo índice cefálico acusa entre 79,5 y 82,5 para ambos padres y que tienen un índice medio de 80,6 tienen hijos cuyo índice medio es de 79,3, esto es 1,3 unidad por debajo del valor de sus padres. Las parejas de bohemios cuyo índice cefálico presenta los mismos límites, y que tienen un índice medio de 81,0 tienen hijos cuyo índice medio es de 83,0 o sea 2 unidades por encima de la de sus padres. Esto demuestra que desde el punto de vista genético los individuos del mismo índice cefálico de estos dos grupos no son idénticos.
Condiciones similares prevalecen respecto de otras características corporales. En otras palabras, individuos de la misma forma corporal pertenecientes a dos poblaciones diferentes son genéticamente no idénticos. Los biólogos han arribado a las mismas conclusiones. Lotry en un cuidadoso análisis del significado de «especie» acentúa la importancia de la identidad constitucional contra la grosera identidad morfológica aparente. La identidad constitucional sólo puede descubrirse por la crianza pura y el cruzamiento, y a menudo se encuentra que formas aparentemente semejantes procrean de distinto modo. De estas observaciones se sigue que debemos describir cada población como un todo, que no debemos separar arbitrariamente a un grupo de cierta apariencia corporal, del resto. El error de identificar a individuos de la misma apariencia corporal, pero pertenecientes a poblaciones diferentes, como miembros de la misma raza es excesivamente común y en él incurren hasta investigadores escrupulosos.
Durante los últimos veinticinco años se han efectuado numerosos estudios de herencia o genética, como se le denomina ahora generalmente, y su resultado puede formularse así: sí el número de vástagos de una pareja única fuera infinitamente grande, entonces la distribución de frecuencia de formas en la prole de esta pareja estaría definidamente determinada por las características orgánicas de los padres, siempre que no hubiera perturbaciones debidas a condiciones externas. Las formas de distribución de frecuencia varían considerablemente, pero para cada pareja son absolutamente fijas, mientras las condiciones externas que influyen sobre la forma corporal continúan siendo las mismas. Esta es la expresión más generalizada de la ley mendeliana de la herencia. Es difícil dar la prueba exacta de estas condiciones en el hombre y en aquellas especies animales en que el número de descendientes es pequeño; pero las observaciones entre animales y plantas inferiores, y la concordancia de las condiciones observadas en el hombre, con las anotadas en formas inferiores son concluyentes. Si bien no podemos decir cuáles pueden ser las características específicas de un individuo cualquiera, el grupo en conjunto siempre se comportará de la misma manera.
Se deduce de estas observaciones que en un estudio estrictamente biológico debemos investigar las líneas genéticas que constituyen una raza antes de poder compenetrarnos del carácter de la raza en su conjunto.
En los animales superiores la progenie es siempre la resultante de la unión de dos individuos, y no conocemos una sola población de animales u hombres en que el macho y la hembra que se unen representen idénticos linajes. Aun en el apareamiento de animales de la misma carnada de hermanos y hermanas la estructura de las células sexuales no es la misma, Las líneas familiares en toda población a pesar de la uniformidad de su origen, son desiguales.
La importancia de estas consideraciones resultará más evidente al analizar la constitución de las poblaciones con mayor detalle.
En una gran población tan poco estable en su hábitat como la de Europa moderna o la moderna Norte América, el número de antepasados de una sola persona aumenta muy rápidamente; siendo dos el número de los padres, cuatro el de los abuelos, ocho el de los bisabuelos, el número teórico de antepasados en veinte generaciones sería de más de un millón, o más precisamente, de 1.048.576. Veinte generaciones representan, conforme al ritmo de crecimiento de los tiempos modernos, alrededor de 700 años; de acuerdo con el ritmo de crecimiento de antaño más o menos 400 años como mínimo. Estas cifras se aplicarían a la serie é generaciones representadas por los primeros hijos varones; para las primeras hijas mujeres las cantidades respectivas serían alrededor de 500 años y 350 años. No obstante si consideramos el verdadero origen de las familias, incluyendo a individuos nacidos más tarde, quizá podríamos suponer que veinte generaciones en Europa representarían de 800 a 900 años y entre los pueblos primitivos quizás un poco menos solamente, toda vez que en tiempos remotos la diferencia entre la rapidez de las generaciones sucesivas en Europa y en los pueblos primitivos no era muy considerable. Esto demuestra que es enteramente imposible que un número tan grande de antepasados como requiere la teoría pueda haber contribuido a la formación de los individuos de la generación actual. La explicación es sencilla. Debido a interalianzas entre las mismas familias un gran número de antepasados serán duplicados en diferentes líneas paternas y maternas; y en esta forma la verdadera ascendencia de cada individuo parece ser mucho más compleja de lo que el tratamiento puramente aritmético sugeriría. El cálculo del cuadro genealógico del ex Emperador de Alemania, por ejemplo, es instructivo. Según O. Lorenz el número de sus antepasados en generaciones sucesivas es el siguiente:
|
Generación |
Número real |
Número teórico |
|
I |
2 |
9 |
|
II |
4 |
4 |
|
III |
8 |
8 |
|
IV |
16 |
14 |
|
V |
32 |
24 |
|
VI |
64 |
44 |
|
VII |
128 |
74 |
|
VIII |
256 |
116* |
|
IX |
512 |
177* |
|
X |
10S4 |
256* |
|
XI |
2048 |
342* |
|
XII |
4096 |
533* |
* Estas generaciones no se conocen completamente. Los valores aquí citados son los valores máximos que podrían encontrarse siempre que los individuos desconocidos no hubieran tenido ninguna «perdida de antepasados».
Una serie de cuarenta familias reales arroja los siguientes promedios:
|
Generación |
Promedio numérico |
|
I |
2,00 |
|
II |
4,00 |
|
III |
7,75 |
|
IV |
13,88 |
|
V |
23,70 |
|
VI |
40,53 |
Cuando comparamos estas condiciones en la inestable población de partes densamente pobladas de Europa y América moderna con las condiciones prevalecientes entre tribus primitivas, resulta obvio que el número total de antepasados de cada individuo en las pequeñas comunidades debe ser mucho menor que el número de antepasados en los estados modernos a que nos acabamos de referir. Un ejemplo característico lo presenta el esquimal de Smith Sound, en Groenlandia del Norte. Por lo que sabemos parece en extremo improbable que esta comunidad estuviera alguna vez compuesta por más de algunos centenares de individuos. El tipo de vida de las comunidades esquimales sugiere que originariamente consistía de unas pocas familias. La comunidad ha permanecido separada del mundo exterior por lapsos muy prolongados; y si bien pueden haber ocurrido advenimientos de nuevos individuos de afuera una que otra vez en cada siglo, en conjunto, ha permanecido aislada. Es por lo tanto indudable que la ascendencia de este grupo no puede contener nada semejante al millón de personas requerido teóricamente sino que todos los individuos deben estar emparentados por medio de sus antepasados inmediatos o remotos.
En una comunidad de este tipo, cuyos miembros nunca han ascendido a más de aproximadamente 200, los antepasados de cada individuo desde la octava generación deben haber sido en su mayor parte los mismos en combinaciones variables, porque la octava generación requeriría, teóricamente, doscientos cincuenta y seis individuos, muchos más de los que en realidad existían en la comunidad. Por lo tanto la existencia de un individuo que no tenga muchos antepasados próximos y remotos en común con todo el resto de la comunidad es muy improbable, si no imposible.
Hemos tratado de determinar la pérdida de antepasados respecto a los bastardos sudafricanos, descendientes de hotentotes y boers. Las tablas genealógicas reunidas por Eugen Fischer [31] dan los siguientes números:
|
Generación |
Nº de antepasados de varias familias |
||
|
Familia I |
Familia II |
Familia III |
|
|
I |
2 |
2 |
2 |
|
II |
4 |
4 |
4 |
|
III |
8 |
8 |
8 |
|
IV |
14,1 |
14,3 |
16 |
|
V |
20,1 |
19,7 |
32 |
|
VI |
32,0 |
— |
|
Estos números son similares a los que se encuentran entre las casas reales de Europa.
Se obtiene un cuadro algo más claro cuando determinamos el número de ascendientes y consideramos cada individuo como miembro de una fraternidad que abraza cierto número de hijos que continúan propagándose con la misma rapidez. Si se trata de una población extensa y móvil podríamos suponer más aún, que los cónyuges de las generaciones sucesivas no están emparentados en ningún grado entre sí o con los miembros del linaje familiar que está en consideración. En tales condiciones el linaje de un individuo cualquiera será aquella fracción del número de antepasados que se obtiene dividiendo el número total de sus antepasados por el número de miembros de su generación. Cuando, por ejemplo, una pareja tiene dos hijos, el promedio de número de padres para cada hijo será 1. Cuando esos hijos se casen y tienen dos hijos, el número total de individuos en la primera generación será de seis, porque los dos hijos de la pareja original tienen los mismos padres. Los cuatro nietos de la pareja original tendrán por lo tanto seis abuelos o sea 1,5 por cada uno. En esta forma se obtiene la siguiente serie de antepasados en línea de descendencia directa.
La aparente contradicción de estos valores —por ejemplo que cuatro nietos tengan seis abuelos— reside en el hecho de que dos de estos nietos son al mismo tiempo, descendientes directos de otra familia. Los parentescos colaterales se extienden rápidamente. Encontramos un cálculo aproximado de estos valores en Jankowsky[10].
Debemos recordar que las condiciones reales dependerán principalmente de la movilidad de la población. Cuando la población es sedentaria y grupos relativamente pequeños están en contacto permanente, un alto grado de crianza sin mezcla con segregación de grupos locales será el resultado, mientras que en una población extensa que se mueve libremente la rapidez con que se desarrolla la crianza será mucho menor.
|
Generación |
Número de antepasados por familia de |
|||
|
2 Hijos |
3 Hijos |
4 Hijos |
5 Hijos |
|
|
I |
1 |
0,67 |
0,50 |
0,40 |
|
II |
1,50 |
0,89 |
0,62 |
0,48 |
|
III |
2,75 |
1,63 |
1,16 |
0,90 |
|
IV |
5,38 |
3,21 |
2,29 |
1,78 |
|
V |
10,69 |
6,40 |
4,57 |
3,55 |
|
VI |
21,34 |
12,80 |
9,14 |
7,11 |
|
VII |
42,67 |
25,60 |
18,29 |
14,22 |
|
VIII |
85,34 |
51,20 |
36,57 |
28,44 |
|
IX |
170,64 |
102,40 |
73,14 |
56,88 |
|
X |
341,33 |
204,80 |
146,28 |
115,78 |
Tanto la crianza sin mezcla de razas como el continuo cruzamiento producen el efecto de que cuando el proceso persiste durante un tiempo prolongado todos los linajes familiares se parecen mucho, mientras que en una población de origen mixto o sin crianza pura los linajes familiares son completamente distintos. Puede ocurrir pues que las formas corporales en dos poblaciones estén distribuidas de la misma manera, si sólo se considera a los individuos, y sin embargo la composición biológica de las dos series sea completamente diferente. En una podemos tener linajes familiares completamente distintos entre sí, siendo todos los hermanos y hermanas de cada familia muy parecidos; en la otra podemos encontrar linajes familiares muy semejantes, mientras que hermanos y hermanas pueden variar mucho entre sí.
El efecto de la crianza pura ha sido probado con animales. Experimentos (King) con ratas, en que hermanos y hermanas fueron apareados por 25 generaciones sucesivas, demuestran que la variabilidad fraternal disminuye gradualmente. Esto parecería indicar tanto una disminución en la variabilidad de los linajes familiares, como en la variabilidad fraternal, pero los datos no nos permiten distinguir entre estos dos caracteres.
Muy pocas poblaciones y sólo contados rasgos han sido examinados desde estos puntos de vista. El material recogido hasta ahora índica que las diferencias entre los linajes familiares que constituyen una población son menores cuanto más estable sea la población y cuanto más tiempo haya durado el proceso de crianza pura, sin selección. Cuando los progenitores de estos linajes familiares son de distinta forma corporal, los hermanos y hermanas de cada familia pueden no parecerse; si los progenitores son semejantes en forma, entonces tanto los linajes familiares como las fraternidades (esto es, hermanos y hermanas de cada familia) serán parecidos.
Los siguientes datos relativos a la variabilidad del valor del ancho de la cabeza expresado en relación al largo de la misma, ilustran las condiciones que se encuentran en varios grupos locales.
Esto significa que entre los armenios cuyo índice cefálico está en el término medio de 85,6, 68% de las familias tienen un índice entre 83,4 y 87,8, mientras que el 32% restante tiene un índice de valores fuera de estos límites. Significa también que 68% de los hermanos y hermanas tienen un índice cefálico que varía entre 3,20 unidades por debajo y por encima del promedio familiar, mientras que el resto está fuera de estos límites. Y ocurre lo mismo con los demás valores.
Las cifras correspondientes a los bastardos son interesantes. Los bastardos son un pueblo descendiente de hotentotes y holandeses que se han casado casi exclusivamente entre ellos durante el último siglo. A pesar de su origen mixto, los linajes familiares son muy semejantes mientras que hermanos y hermanas presentan variaciones considerables. Las condiciones entre los indios chippewa de Canadá y particularmente entre los missisauga, rama local de los chippewa, son muy semejantes. Éstos constituyen una antigua población mixta de descendientes de indios, franceses e irlandeses. Entre los negros americanos encontramos también mayor uniformidad de linajes familiares porque ellos representan también una antigua mezcla de blancos y negros.
| Variabilidad[11] normal del índice cefálico | ||||
|
I Total |
II Fraternal |
III Linajes familiares |
Razón de II y III |
|
|
Armenios |
3,88 |
3,20 |
2,20 |
1,46 |
|
Indios Chippewa |
3,76 |
3,32 |
1,77 |
1,88 |
|
Italianos del centro |
3,62 |
2,72 |
2,39 |
1,14 |
|
Negro-blancos de Nueva York |
3,51 |
2,93 |
1,85 |
1,58 |
|
Bohemios |
3,53 |
2,61 |
2,37 |
1,10 |
|
Escoceses |
3,43 |
2,66 |
2,17 |
1,21 |
|
Missisauga |
3,43 |
3,10 |
1,47 |
2,11 |
|
Judíos del Este de Europa |
3,40 |
2,52 |
2,29 |
1,10 |
|
Worcester Massachusetts |
3,34 |
2,36 |
2,36 |
1,00 |
|
Holandeses |
3,05 |
2,33 |
1,95 |
1,20 |
|
Bastardos de Sur África |
2,82 |
2,52 |
1,26 |
2,00 |
|
Blancos de la Montaña Blue Ridge |
2,80 |
2,09 |
1,85 |
1,15 |
El significado de estas cifras puede ilustrarse más acabadamente por medio de la siguiente consideración: en la población missisauga un 16% de las familias tienen un índice superior a 1,47 unidad por debajo del término medio.
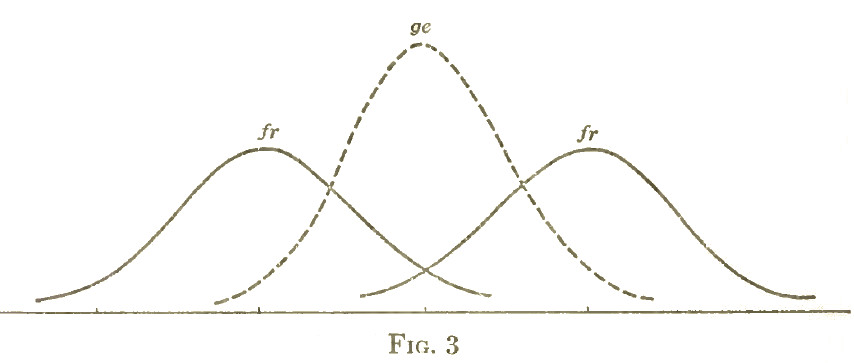
Toda vez que la variabilidad fraternal es ± 3,10, o sea más de dos veces la variabilidad de los linajes familiares, habrá una superposición considerable entre estos dos grupos (fig. 3).
De acuerdo con las constantes de las estadísticas, alrededor del 32% del grupo más bajo tendrá valores por encima y el mismo porcentaje del grupo más alto tendrá valores por deba) o del término medio general, de modo que alrededor de un 32% de cada uno de los 16%, o sen alrededor del 10% de los extremos de la población tendrá las mismas formas. En Worcester en cambio, sólo 16% de los dos extremos se superpondrán, así que solamente el 5% de los extremos se superpondrán.
fr= Variabilidad de fraternidades
ge= Variabilidad de linajes familiares
Esto es más claro cuando los extremos elegidos están a mayor distancia del término medio. Podríamos considerar el grupo de los missisauga, que están alejados más de 2,2 unidades del término medio. Hay un 6,7% de la serie total por debajo y 6,7% por encima de estos puntos. Luego, de acuerdo con las estadísticas de las constantes alrededor del 24% de cada uno de estos grupos o sea 3,2% de la población total se superpondrá. Para la población de Worcester sólo 6,7% de cada uno de los grupos extremos se superpondrá, de modo que menos del 1% será común a las dos series[12].
Es necesario tener esto presente pues algunos autores, como Fritz Lenz, subestiman la significación de las diferencias genéticas dentro de la raza.
Debe recordarse que esta discusión se refiere a un solo rasgo. Si los rasgos estudiados son más numerosos, la heterogeneidad de las familias se tomará aún más manifiesta.
Deducimos de ello que en la mayoría de las poblaciones los linajes familiares difieren tanto que siempre se hallarán líneas distintivas. En contraste con esto, es imposible encontrar distinciones radicales análogas entre poblaciones completas de superficies contiguas.
Así pues, si bien es imposible dar una definición biológica exacta de una raza, podemos definir los linajes familiares con mucho mayor precisión y por lo tanto la raza debe ser definida como un complejo de linajes familiares. El origen y carácter de los linajes familiares determina el carácter de la raza.
Los resultados de nuestro estudio están en completo acuerdo con los puntos de vista modernos respecto a lo que constituye una especie de animales o plantas. La disolución que Johannsen realiza de la especie o como él lo denomina, el fenotipo, en una serie de genotipos, es comparable a nuestro análisis de una raza. Johannsen estudió principalmente habas autofecundadas. En este caso el fenómeno es por supuesto mucho más claro de lo que puede ser en casos en que la autofecundación es imposible y donde el cruzamiento de los linajes ocurre constantemente. El punto de vista aquí expresado concuerda también con el de O. F. Cook quien reconocía solamente al individuo y sus descendientes en las especies. Él también considera a la especie un complejo de líneas distintas.
Cuanto más gradual es la transición entre los tipos locales, tanto más necesario es tener bien presente este punto. Podemos llamar heterogénea a una población en que los linajes familiares son muy diferentes porque un solo linaje familiar no es representativo de toda la población. En este sentido las poblaciones con baja variabilidad de linajes familiares pueden llamarse homogéneas. Puede muy bien ocurrir que en tal caso los antepasados sean completamente diversos, como entre los bastardos sudafricanos, descendientes de holandeses y hotentotes, y sin embargo las familias pueden ser tan semejantes que cada una represente justa y adecuadamente el tipo general de la población entera.
Nunca puede hallarse una homogeneidad absoluta debido a las leyes de la herencia, por más puro que sea el cruzamiento ocurrido. Mientras no se lleven a cabo investigaciones más extensas acerca de esta cuestión, no estaremos en condiciones de decir cuál puede ser el límite de homogeneidad de una población.
Aun sin información detallada es fácil ver que el grado de homogeneidad debe variar considerablemente. Los habitantes de una pequeña y estable aldea europea en que la tierra permaneció en poder de las mismas familias durante siglos deben tener un alto grado de homogeneidad. Ocurrirá lo mismo en pequeñas tribus aisladas.
Las grandes ciudades representan las condiciones inversas. Debido a la confluencia de gente de distinto origen, los linajes familiares serán sumamente variados. El carácter de las fraternidades y la variabilidad de los linajes familiares experimentan constantes cambios a medida que prosigue la integración de la población, hasta que finalmente se establece una nueva condición estable, siempre que no ocurra ningún nuevo aumento de linajes extraños, condición que en la vida de la ciudad nunca se realiza.
La composición de una raza puede también ser observada desde otro ángulo, Cuando comparamos dos tipos completamente distintos, todos los individuos de cada tipo nos parecen iguales y diferentes del otro tipo. Por el contrario cuando dos tipos se superponen, las diferencias individuales resultarán más notables y el grado de semejanza mutua entre los miembros de cada tipo no parecerá tan grande. Cuando comparamos una familia negra con una familia blanca, las pequeñas diferencias entre los hermanos y hermanas de cada una de esas familias parecen insignificantes. Cuando comparamos en cambio dos familias que son muy semejantes, las diferencias individuales parecerán mucho más importantes y por tanto las semejanzas familiares entre hermanos y hermanas parecerán muy leves. En una sola familia, si no la comparamos con ninguna otra familia, los hermanos y hermanas son simplemente diferentes. Sólo individualmente tienen grados diferentes de semejanza. Otro tanto ocurre cuando tenemos un número de familias idénticas: todos los individuos serán diferentes y no habrá parecidos familiares. Esto no altera el hecho de que la variabilidad fraternal en las familias pueda ser grande o pequeña.
Esta sencilla consideración demuestra que la semejanza fraterna depende totalmente de la composición de la población. En una población muy heterogénea las semejanzas fraternas serán grandes; en una homogénea serán pequeñas. Lo mismo puede decirse de las semejanzas entre padres e hijos. Nos parecerá tanto más notable cuanto mayor sea la heterogeneidad de la población, mientras que en una población comparativamente homogénea desaparecerá prácticamente, porque en cada familia estará representado el mismo tipo. Cuando Francis Galton [3] estudió este fenómeno destacó la importancia del grado de semejanza entre hermanos y hermanas y padres e hijos, lo que él expresaba por el llamado coeficiente de regresión. Lo determinaba del siguiente modo: cuando padres o madres, en una población dada, difieren en cierta medida del término medio general de la población, los niños diferirán de la población general en cierta fracción de esta medida; y si un individuo difiere en cierta medida del término medio de la población, sus hermanos o hermanas diferirán, en general, del término medio de la población, en una cierta fracción de esta medida.
Para las poblaciones a que nos referimos en la pág. 74 y para las desviaciones medias del índice cefálico de hermanos y hermanas se han hallado los siguientes valores de estas fracciones:
|
Worcester, Massachusetts |
0,50 |
|
Bohemios |
0,45 |
|
Judíos del Este de Europa |
0,45 |
|
Blancos del Blue Ridge |
0,44 |
|
Italianos del Centro |
0,44 |
|
Holandeses |
0,41 |
|
Escoceses |
0,40 |
|
Armenios |
0,30 |
|
Negro-blancos de Nueva York |
0,28 |
|
Indios Chippewa |
0,21 |
|
Bastardos Sud Africanos |
0,20 |
|
Missisauga |
0,18 |
Por ejemplo, si en Worcester, Massachusetts, un individuo tiene un índice cefálico de 4 unidades sobre el término medio, sus hermanos y hermanas tendrían, término medio, un índice de 4 x 0.5 o sea 2 unidades sobre el término medio; mientras que entre los missisauga los hermanos y hermanas tendrían, término medio, un índice cefálico de sólo 4 x 0,18 o sea 0,72 sobre el término medio. Las diferencias en estos valores se deben a los variables grados de heterogeneidad de la población. Las series más homogéneas tienen las correlaciones más bajas. La relación de acuerdo a la variabilidad de los linajes familiares (I) y la razón entre el linaje familiar y la variabilidad fraternal (II) es la siguiente:
|
I |
II |
||
|
Bastardos |
± 1,26 |
Missisauga |
2,11 |
|
Missisauga |
± 1,47 |
Bastardos |
2,00 |
|
Chippewa |
± 1,77 |
Chippewa |
1,88 |
|
Negro-blancos de N. York |
± 1,85 |
Negro-blancos de N. York |
1,58 |
|
Blancos del Blue Ridge |
± 1,85 |
Armenios |
1,46 |
|
Holandeses |
± 1,95 |
Escoceses |
1,21 |
|
Escoceses |
± 2,17 |
Holandeses |
1,20 |
|
Armenios |
± 2,20 |
Blancos del Blue Ridge |
1,13 |
|
Judíos del E. de Europa |
± 2,29 |
Italianos del centro |
1,14 |
|
Worcester, Massachusetts |
± 2,36 |
Judíos europeos del Este |
1,10 |
|
Bohemios |
± 2,37 |
Bohemios |
1,10 |
|
Italianos del centro |
± 2,39 |
Worcester, Massachusetts |
1,00 |
El acuerdo entre estas dos columnas demuestra que en las series elegidas, la razón de la variabilidad de las fraternidades medida por la variabilidad de los linajes familiares es mayor cuanto más uniformes son los linajes familiares.
Debemos volver una vez más a la discusión de los linajes familiares.
Suponíamos en las consideraciones previas que todas las familias de una población tendrán la misma variabilidad fraterna. Toda vez que el origen de los linajes familiares no es uniforme, esto no es probable y los valores que citamos antes, deben ser considerados como aproximaciones a las condiciones reales.
Puede demostrarse que dentro de la misma población, la variabilidad de fraternidades aumenta con la diferencia entre los padres. El estudio de un considerable número de familias demuestra que la variabilidad del índice cefálico dentro de una población aumenta con la diferencia de índice cefálico entre los padres (Boas 5).
|
Diferencia de índice cefálico entre los padres |
Variabilidad fraternal Nueva York |
Casos |
Variabilidad fraternal, Holanda |
Casos |
|
0-2,9 unidades |
6,8 |
1102 |
5,3 |
627 |
|
3.5,9 unidades |
6,7 |
756 |
5,9 |
473 |
|
6-8,9 unidades |
8,5 |
317 |
5,4 |
182 |
|
9 y más unidades |
13,0 |
108 |
8,5 |
66 |
Félix von Luschan [1] halló un fenómeno similar en la población mixta del sud de Asia Menor donde un pueblo de cabeza redonda del Asia Central se ha mezclado durante miles de años con el pueblo de cabeza alargada de la costa de Siria. Aquí también encontramos un considerable aumento en la variabilidad de la población mixta si se la compara con el grado de variabilidad encontrado en poblaciones más homogéneas.
La distribución de las formas de las cabezas en Italia también ilustra este punto. En Italia central, donde los italianos del norte, de cabeza ancha, y los italianos del sur, de cabeza alta han contraído alianzas, la variabilidad de la forma de la cabeza es grande.
Estos fenómenos son expresiones de las varias formas de herencia mendeliana a que antes nos hemos referido (pág. 67), a la tendencia de los individuos de origen mixto a revertir en razones numéricas definidas, respecto a varias características corporales, a los tipos de los cuales descienden.
A menudo se compara a las razas humanas con las razas modernas de animales domésticos. Existe sin embargo una diferencia fundamental. Las razas de animales domésticos se crían por medio de una selección cuidadosa y la división mendeliana de las familias es eliminada por la cría de sólo aquellos individuos de raza legítima. La variación en una cría de animales domésticos, es, por lo tanto, muy pequeña, tanto más cuanto más cuidadosamente se eliminen todos los tipos mixtos. En esta forma se han desarrollado razas extraordinariamente diferenciadas. Nada semejante ha ocurrido con el hombre. Por el contrario, aún cuando se procuró impedir las alianzas entre diferentes castas, la tentativa no tuvo nunca éxito. Las barreras sociales son destruidas y gradualmente los dos tipos de la población se mezclan entre sí. Esto es cierto aún en tan rígidos sistemas de casta como los de la India. Fue igualmente cierto entre patricios y plebeyos en Roma y a pesar del recrudecimiento de la violenta oposición a la mezcla de razas en Alemania, la historia de la humanidad no ha de ser revocada.
Tipos locales más o menos especializados se desarrollan únicamente cuando pequeños grupos viven aislados y el corto número de antepasados demuestra peculiaridades. Cuanto más pequeño el grupo de antepasados, mayor será la probabilidad de que el grupo local parezca mas o menos distinto del tipo de la población general del que descendió. Cuando encontramos, por ejemplo, en Norte América un tipo muy marcado en la costa ártica del continente y otro tipo en la cuenca del Mackenzie, otros aún en localidades bien definidas de la costa del Pacífico, en el valle del Misisipi, en el sudeste, a lo largo del Río Grande y en México, parece posible atribuir su origen al crecimiento de pequeños grupos aislados. Otros casos de la aparición de formas peculiares en comunidades locales pueden explicarse de esta misma manera: por ejemplo la frecuencia desacostumbrada de los Incae (la división del hueso occipital por una sutura transversal) en Perú y los indios pueblo (Mathews 1) y la gran frecuencia del torus palatinas (arruga a lo largo de la línea media del paladar) entre los lapones y en las costas orientales del Báltico (Lissauer).
Puede señalarse aquí otro fenómeno que se ha investigado poco aún pero merece atenta observación. Hemos visto que en comunidades estables, en países de poblaciones dispersas, el parentesco entre los miembros de un grupo será muy estrecho y este parentesco afectará necesariamente el tipo y su variabilidad. En el curso del tiempo, dos territorios cuyas poblaciones se han desarrollado así pueden entrar en contacto y pueden ocurrir numerosos matrimonios entre ellos. Se verá en seguida que aunque las diferencias entre los dos tipos puedan ser aparentemente muy leves, el resultado será un completo trastorno en las formas de herencia, porque un gran número de individuos de distinto origen se han puesto en contacto. Por ejemplo, los italianos del sur y los españoles representan dos tipos no muy distintos en rasgos físicos, pero separados por siglos. Las pequeñas comunidades de las aldeas de Italia, lo mismo que las de España, tienen todas las características de comunidades en que los matrimonios endogámicos han ocurrido durante un largo período. En la República Argentina estos dos tipos entran en contacto y se casan unos con otros con frecuencia. No tenemos observaciones acerca del efecto de esta mezcla sobre las características físicas, pero se ha observado que la distribución de nacimientos masculinos y femeninos es completamente diferente a la que prevalece en familias en que ambos padres son españoles o italianos (Pearl 3). Es también concebible que esto pueda ser uno de los elementos que producen el cambio de tipo de las poblaciones urbanas en comparación con las poblaciones rurales en Europa y que puede haber tenido activa participación en el cambio de tipo observado entre los descendientes de inmigrantes europeos en América.
Todos los intentos de reconstruir los elementos componentes de una población de origen mixto están destinados a fracasar. Suponiendo, por ejemplo, que no conociéramos una raza blanca y una raza negra, sino solamente a los mulatos ¿podríamos reconstruir la raza blanca y la raza negra? Si conociéramos las leyes de herencia de cada rasgo individual, sus interrelaciones y los cambios que pueden ocurrir debido a la mezcla; si además supiéramos cuáles han sido las influencias del medio ambiente y la selección, esto podría parecer posible; pero dichos intrincados mecanismos son conocidos muy imperfectamente, y la tarea sería semejante a la de una persona que ha de resolver una ecuación simple con muchas cantidades desconocidas y casi sin guía alguna en la elección de las cantidades que llenarían las condiciones de la ecuación original.
Esto es aun más cierto en tipos que son semejantes como los de Europa y el Cercano Oriente, cuyas características corporales son tan poco divergentes que no se puede asignar a los individuos con certeza a uno u otro grupo. Todo lo que sabemos es que cada grupo consiste de muchos linajes familiares divergentes. No es posible efectuar la reconstrucción de los linajes «puros» de los que deriva la población actual. Pueden darse infinidad de soluciones estadísticas, pero su interpretación biológica requerirá un conocimiento cabal de las condiciones que gobiernan los efectos de la mezcla de distintos linajes familiares.
No podemos predecir ni siquiera estadísticamente, mucho menos individualmente, cuál será el resultado de la inter-alianza de dos razas. Mucho menos estamos capacitados para invertir el proceso y determinar los tipos de que pueda haber surgido una población.
Hemos discutido hasta aquí los tipos raciales tal como aparecen en el adulto. Debemos considerar la forma en que las características hereditarias hallan expresión en la evolución del individuo. Las características raciales específicas —esto es, los rasgos cuya variación individual es pequeña comparada con las diferencias raciales—, se establecen, generalmente, a muy temprana edad. A. Schultz ha demostrado que los rasgos característicos del negro y el blanco son perceptibles en la vida fetal. Cuanto más pronunciada la diferencia entre dos tipos, tanto más temprano se establece. Durante este período la diferenciación de tipos raciales es más marcada que la diferenciación individual. En el curso del crecimiento tanto el carácter racial como el individual aparecen más y más acentuados y este proceso continúa a través de la vida entera. Por esta razón las características de los tipos locales son a menudo más distintas en los adultos varones que tienen un período más intenso y prolongado de desarrollo que en las mujeres. Son menos marcadas en los niños. Los tipos raciales más generalizados se encontrarán entre los niños, los tipos más especializados entre los adultos varones. Esto puede ilustrarse por el análisis de la forma de la nariz de niños indios, chinos y blancos que es mucho más semejante que entre los adultos. El caballete de la nariz es bajo, su elevación sobre la cara escasa, el párpado superior tiene a menudo un pliegue interior que confiere al ojo una aparente posición oblicua. Esto se observa particularmente en los mongólicos e indios pero también con mucha frecuencia en niños blancos. Desaparece con el aumento de la elevación de la nariz sobre el plano del rostro. Las proporciones de las extremidades y el cuerpo en los niños de estas tres razas no difieren mucho.
Acompañando la diferenciación de tipos encontramos también una diferenciación de características individuales. Después de llegar a cierta edad la proporción de diferenciación individual sobrepasa a la de diferenciación racial. Las características raciales que no han quedado establecidas antes de ese momento, no se desarrollan en los años posteriores. La edad en que ocurre un marcado progreso en la individualización no es la misma para todos los rasgos. La pigmentación se estabiliza poco después del nacimiento. La forma de la cabeza se estabiliza a la edad de uno o dos años. El desarrollo más típico de la nariz ocurre durante la adolescencia.
Podemos expresar esto en otra forma. Los tipos más generalizados se encuentran en los individuos más jóvenes. El proceso de especialización ocurre durante la infancia y el más alto grado de especialización se encuentra entre los hombres adultos. Las afinidades entre grupos raciales lejanamente emparentados pueden descubrirse, por lo tanto, más fácilmente comparando formas fetales y de niños pequeños. Es casi imposible en la actualidad hacer afirmaciones definidas respecto a esté punto porque se conoce demasiado poco acerca de las formas anatómicas de los niños entre los australianos, bosquimanos, negritos o indios americanos, y éstos se cuentan entre los más importantes de los grupos cuya posición ha de ser determinada.
Parece probable que los diversos grupos tengan cada uno una rapidez característica de desarrollo para los diversos rasgos físicos. No se sabe con certeza si en tales casos las condiciones ambientales desempeñan un papel importante o si nos encontramos ante rasgos hereditarios. La comparación de niños judíos y no judíos que asisten a diferentes escuelas demuestra que los niños judíos crecen al principio más rápidamente que los niños no judíos, mientras que más tarde, el ritmo del crecimiento de estos últimos es más rápido que el de los niños judíos. Bajo iguales condiciones sociales no ocurriría tal diferencia. El orden de aparición de dientes permanentes entre los negros sudafricanos y blancos americanos no es el mismo. Las diferencias observadas entre estos dos grupos es mucho menor que las notadas en linajes familiares donde hay clara certeza de tendencias hereditarias respecto al tamaño y al tiempo de su crecimiento.