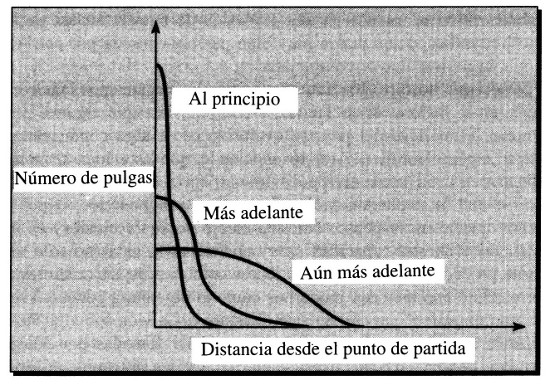
Figura 18. Distribución cambiante de las distancias en un paseo aleatorio
La selección en la evolución biológica y otros ámbitos
Los sistemas complejos adaptativos de todo tipo, incluida la evolución biológica, operan de acuerdo con la segunda ley de la termodinámica. Ahora bien, los antievolucionistas aducen de vez en cuando que la evolución biológica, de la que emergen formas cada vez más complejas, representa un incremento temporal del orden que está en clara contradicción con esta segunda ley. Es un argumento erróneo por diversas razones.
Primero, como ha sido descrito con anterioridad, también en la evolución de sistemas no adaptativos como galaxias, estrellas, planetas y rocas emergen formas cuya complejidad aumenta con el tiempo, sin que eso entre en conflicto con el incremento de entropía. Las estructuras que surgen envejecen de acuerdo con la segunda ley, pero la distribución de complejidad se amplía con el tiempo, con un incremento gradual de la complejidad máxima.
Segundo, la segunda ley de la termodinámica se aplica únicamente a sistemas cerrados (esto es, completamente autocontenidos). Quienes pretenden ver una contradicción entre la segunda ley y la evolución biológica cometen el error crucial de fijarse sólo en los organismos sin tener en cuenta el entorno.
La razón más obvia que impide considerar a los sistemas vivos como sistemas cerrados es la necesidad de luz solar como fuente de energía directa o indirecta. Estrictamente hablando, no podemos esperar que se cumpla la segunda ley de la termodinámica a menos que se tenga en cuenta la absorción de energía solar. Por otra parte, la energía fluye tanto hacia dentro como hacia fuera del sistema, perdiéndose al final en forma de radiación (piénsese en la radiación térmica emitida desde nuestras casas hacia el frío y oscuro cielo nocturno). El flujo de energía a través de un sistema puede generar orden localmente.
Además de este efecto, hay que tener en cuenta la influencia de la información procedente del medio ambiente terrestre. Para ver lo que ocurre cuando se incluye dicha información, considérese un caso extremadamente simple en que la influencia ejercida por el ambiente es estable y se ignora la interacción entre las diferentes clases de organismos. Se deja entonces evolucionar una población de un determinado organismo en un medio ambiente constante. Con el tiempo, la población tiende a adaptarse mejor a su entorno a través de la competencia entre los diversos genotipos presentes, unos con más éxito que otros en la creación de fenotipos que sobreviven y se reproducen. En consecuencia, una cierta discrepancia informacional entre el entorno y el organismo se va reduciendo gradualmente, un proceso que recuerda el modo en que las temperaturas de un objeto frío y otro caliente en contacto mutuo se aproximan al equilibrio térmico, de conformidad con la segunda ley de la termodinámica. La evolución biológica, lejos de contradecir dicha ley, proporciona metáforas instructivas de la misma. El proceso de adaptación es en sí mismo una especie de envejecimiento de la población dentro de su medio ambiente.
En los manantiales calientes sulfurosos de todo el mundo (y en las profundidades oceánicas donde los surtidores de agua caliente marcan los límites entre placas tectónicas) proliferan organismos primitivos denominados extremófilos (o crenarqueotas) en un entorno que la mayoría de seres vivos encontraría extremadamente hostil. En la vida de los extremófilos del fondo oceánico la luz juega un papel reducido, limitado principalmente a procesos que suministran productos químicos oxidados. La luz, por ejemplo, mantiene otras formas de vida cerca de la superficie del agua, desde la que cae continuamente materia orgánica hacia el hábitat de los extremófilos.
Hay evidencias indirectas que sugieren con fuerza la existencia, hace más de tres mil millones de años, de organismos que eran similares, por lo menos desde el punto de vista metabólico, a los modernos extremófilos. Nadie sabe si los genotipos de aquellos organismos eran también similares o si parte del genoma habría experimentado una deriva genética sustancial sin afectar de modo significativo a los resultados de las presiones selectivas en el mundo real. En cualquier caso, se puede decir que el difícil problema de vivir en ambientes calientes, ácidos y sulfurosos fue resuelto cuando la Tierra era joven. Los extremófilos alcanzaron una especie de estado estacionario, algo parecido a un equilibrio evolutivo con sus alrededores.
Pero raramente el medio ambiente es tan estable. La mayoría de situaciones naturales son más dinámicas, con cambios medioambientales significativos a lo largo del tiempo. Un ejemplo es la composición de la atmósfera de la Tierra, que en gran medida se debe a la presencia de la vida. La existencia de cantidades significativas de oxígeno en la actualidad puede atribuirse, al menos en gran parte, a las plantas que han proliferado en la superficie del planeta en el pasado.
El medio ambiente de los organismos de cualquier especie incluye un número enorme de otras especies que evolucionan a su vez. El genotipo de cada organismo, o bien el conjunto de genotipos que caracteriza a cada especie, puede contemplarse como un esquema que incluye una descripción de muchas de las otras especies junto con sus reacciones probables ante diferentes formas de comportamiento. Una comunidad ecológica consiste entonces en un gran número de especies que desarrollan modelos de los hábitos de las otras especies y de cómo tratar con ellas.
En algunos casos es una idealización útil considerar sólo dos especies en coevolución, respondiendo cada una a los progresos de la otra. Por ejemplo, en mis caminatas por las selvas de Sudamérica me he encontrado a menudo con una especie de árbol que suministra alimento a una especie de hormiga particularmente agresiva. A cambio la hormiga repele a muchos animales, humanos incluidos, que podrían dañar el árbol. Igual que yo he aprendido a reconocer estos árboles para evitar tropezar accidentalmente con ellos, otros mamíferos han aprendido a evitar ramonear en ellos. Esta simbiosis tiene que ser fruto de un sustancial periodo de coevolución.
En estas mismas selvas pueden observarse relaciones ofensivo-defensivas fruto también de la evolución de dos especies adaptándose mutuamente. Un árbol puede desarrollar la capacidad de exudar una sustancia tóxica capaz de repeler algún insecto destructivo. El insecto puede, a su vez, desarrollar la capacidad de metabolizar el veneno, que deja así de constituir un peligro. La evolución posterior del árbol puede tener como resultado una modificación del veneno que lo haga activo de nuevo, y así sucesivamente. Tales carreras de armamentos químicos desembocan muchas veces en la producción de agentes bioquímicos muy potentes. Algunos de ellos pueden ser muy útiles en medicina, control de plagas y otros campos.
En las situaciones reales no simplificadas muchas especies evolucionan juntas en el seno de una comunidad ecológica, con un entorno no vivo que se altera gradualmente (o incluso rápidamente) con el tiempo. Esto está lejos de los ejemplos idealizados de simbiosis o competencia entre dos especies, así como éstos son más complicados que el caso aún más ideal de una sola especie evolucionando en un medio ambiente inmutable. En todos los casos el proceso evolutivo es compatible con la flecha del tiempo termodinámica, siempre que se tenga en cuenta la totalidad del sistema; pero sólo en las situaciones más simples, como es el caso de los extremófilos, la evolución conduce a una especie de estado estacionario informacional. En general, se trata de un proceso de cambio dinámico continuo, semejante a los que tienen lugar en el seno de sistemas fisicoquímicos complejos como una galaxia, una estrella o un planeta carente de vida. Todos envejecen y decaen con el paso del tiempo, aunque de una manera complicada.
En una comunidad ecológica, el proceso de adaptación mutua a través de la evolución es un aspecto de este envejecimiento. La evolución biológica es parte del proceso de decaimiento que tiende a reducir el intervalo informacional entre las tendencias potenciales y las reales. Una vez existe un sistema complejo adaptativo, el descubrimiento y explotación de oportunidades es no sólo posible, sino probable, porque el sistema es empujado en esta dirección por las presiones selectivas que operan sobre él.
La evolución biológica no suele proceder a un ritmo más o menos uniforme, como imaginaban algunos especialistas. En vez de eso tiende a exhibir lo que se llama un «equilibrio puntuado», en el que las especies (así como los grupos taxonómicos de orden superior: géneros, familias y demás) permanecen relativamente inmutables, al menos en lo que respecta al fenotipo, durante largos periodos de tiempo y después experimentan cambios comparativamente rápidos en un breve lapso de tiempo. Stephen Jay Gould, que propuso esta idea en diversos artículos técnicos escritos en colaboración con Niles Eldredge, ha publicado una extensa literatura sobre el tema en sus amenos artículos y libros populares.
¿Cuál es la causa de los cambios comparativamente rápidos que constituyen las puntuaciones? Los mecanismos posibles pueden dividirse en varias categorías. Una comprende alteraciones, a veces generalizadas, en el entorno fisicoquímico. Hacia el fin del periodo Cretácico, hace unos 65 millones de años, al menos un objeto de gran tamaño, el que formó el enorme cráter de Chicxulub en la península del Yucatán, colisionó con la Tierra. Los cambios atmosféricos resultantes contribuyeron a la extinción del Cretácico, en la que desaparecieron los dinosaurios y muchas otras formas de vida. Cientos de millones de años antes, durante el periodo Cámbrico, se abrieron multitud de nichos ecológicos que fueron ocupados por nuevas formas de vida (algo parecido al modo en que la aparición de una tecnología nueva y popular crea numerosos puestos de trabajo). Estas formas de vida propiciaron a su vez la aparición de nuevos nichos, y así sucesivamente. Algunos teóricos de la evolución conectan esta explosión de diversidad con un incremento en el contenido de oxígeno de la atmósfera, pero esta hipótesis no es aceptada todavía por todo el mundo.
Otra categoría de cambio rápido que puede puntuar el aparentemente estacionario equilibrio evolutivo tiene un carácter eminentemente biológico, y no requiere cambios repentinos importantes en el entorno físico. Es el resultado de la tendencia del genoma a experimentar cambios graduales que no afectan profundamente la viabilidad del fenotipo. Como resultado de este proceso de «deriva», el conglomerado de genotipos que constituye una especie puede desplazarse hacia una situación inestable en la que cambios genéticos ínfimos pueden alterar radicalmente el fenotipo. Puede ocurrir que en un momento dado cierto número de especies de entre las que constituyen una comunidad se estén aproximando a este tipo de inestabilidad, creando una situación propicia para la ocurrencia de mutaciones que conduzcan a importantes cambios fenotípicos en uno o más organismos. Estos cambios pueden iniciar una sucesión de acontecimientos en que algunos organismos tienen más éxito y otros mueren, la totalidad de la comunidad se altera y se abren nuevos nichos ecológicos. Tales trastornos pueden provocar cambios en las comunidades vecinas a través de, por ejemplo, la migración de nuevas especies de animales que entran en competencia con las residentes. Se ha producido una puntuación en el seno de un equilibrio temporal aparente.
Determinados hechos biológicos especialmente cruciales son a veces responsables de ejemplos sobresalientes de equilibrio puntuado en ausencia de cambios radicales en el entorno fisicoquímico. Harold Morowitz, que trabaja en la Universidad George Mason y en el Instituto de Santa Fe, señala la gran importancia de las rupturas o sucesos umbral que abren nuevos abanicos de posibilidades, a veces relacionadas con funciones o niveles de organización superiores. Harold ha resaltado especialmente los casos en que estos umbrales son únicos o casi, o bien dependen de una innovación bioquímica.
Para comenzar, teoriza sobre los posibles umbrales químicos en el curso de la evolución prebiótica que condujo al origen de la vida sobre la Tierra. Estos umbrales incluyen:
En todos estos casos Harold destaca la estrechez del umbral. Sólo unas pocas reacciones químicas especiales hacen posible la entrada en un nuevo dominio, y a veces es una sola reacción la responsable. (La especificidad de tales reacciones no significa que sean improbables. Incluso una reacción única puede tener lugar fácilmente.)
En la evolución biológica tuvieron lugar sucesos umbral análogos que condujeron al desarrollo de todas las formas de vida actuales a partir de la forma ancestral. Muchos de estos sucesos dieron origen a nuevos niveles de organización. Un ejemplo es la evolución de los eucariotas, organismos cuyas células poseen un núcleo verdadero (que contiene la fracción principal del material genético) y también otros «orgánulos» (mitocondrias o cloroplastos). La transformación de organismos más primitivos en eucariotas unicelulares es para muchos investigadores fruto de la incorporación de otros organismos que se convirtieron en endosimbiontes (es decir, que vivían dentro de la célula y en simbiosis con ella) y evolucionaron después hasta convertirse en orgánulos.
Otro ejemplo es la evolución de los eucariotas unicelulares animales (presumiblemente los ancestros de los animales propiamente dichos). Se piensa que los eucariotas vegetales fotosintéticos aparecieron antes, provistos de una pared celular de celulosa además de una membrana interior (la aparición de membranas requirió otro evento bioquímico, la formación de esteroles, compuestos relacionados con el colesterol y las hormonas sexuales humanas). La evolución condujo después a organismos desprovistos de pared celular que podían dedicarse a devorar organismos fotosintéticos en lugar de efectuar la fotosíntesis por sí mismos. Esta innovación fue la clave para la posterior aparición de los animales propiamente dichos.
La evolución de organismos pluricelulares a partir de organismos unicelulares, presumiblemente a través de la agregación, fue posible gracias a otra innovación bioquímica, un adhesivo que mantenía unidas las células.
Harold Morowitz y otros piensan que, al menos en muchos casos, un pequeño cambio en el genoma —producto de una o a lo sumo unas pocas mutaciones, pero culminando una larga serie de cambios anteriores— puede disparar un suceso umbral e iniciar así una de las revoluciones que representan una puntuación capital dentro de la relativa estabilidad del equilibrio evolutivo. Una vez dentro del dominio abierto por el evento, el organismo adquiere regularidades nuevas y significativas que lo elevan a un nivel más alto de complejidad.
Como ocurre con las perturbaciones físicas de gran magnitud, como un terremoto (o la colisión de la Tierra con otros objetos dentro del sistema solar), tales acontecimientos capitales pueden contemplarse como eventos individuales de gran trascendencia, o bien como sucesos inusuales de gran magnitud en la cola de una distribución que comprende eventos de mucha menor magnitud en su mayoría.
En la evolución de una comunidad ecológica, un sistema económico o una sociedad, igual que en la evolución biológica, se van presentando ocasiones para un incremento de la complejidad. El resultado es que la complejidad máxima tiende a aumentar. Los incrementos de complejidad más fascinantes son los que tienen que ver con una transición a un nivel de organización superior, típicamente a través de la formación de agregados, como en la evolución de los animales y vegetales pluricelulares a partir de organismos unicelulares.
Familias o bandas de seres humanos pueden agruparse para formar una tribu. Un grupo de personas puede aunar sus esfuerzos para ganarse la vida montando una empresa. En el año 1291, tres cantones, a los que pronto se unió un cuarto, formaron una confederación que con el tiempo se convertiría en la moderna Suiza. Las trece colonias norteamericanas se unieron en una confederación que más tarde, ratificando la constitución de 1787, se convirtió en una república federal llamada Estados Unidos. La cooperación conducente a la agregación puede ser efectiva.
Aunque la competencia entre esquemas es una característica de los sistemas complejos adaptativos, los sistemas mismos pueden exhibir una mezcla de competencia y cooperación en sus interacciones mutuas. A menudo resulta beneficioso para estos sistemas unirse formando una entidad colectiva que funciona también como un sistema complejo adaptativo, por ejemplo cuando en un sistema económico personas y empresas operan bajo las normas de un gobierno que regula su comportamiento para promover valores importantes para el conjunto de la comunidad.
También entre esquemas en competencia es posible a veces que se establezca una cooperación ventajosa. En el dominio de las teorías, por ejemplo, las nociones en competencia no siempre son mutuamente excluyentes. A veces la síntesis de varias ideas está más cerca de la verdad que cualquiera de ellas por separado. Pero quienes proponen una aproximación teórica particular saben que es más fácil obtener recompensas en la vida académica y en otros ámbitos pretendiendo que la propuesta propia es absolutamente correcta y enteramente nueva, y arguyendo que los puntos de vista alternativos son erróneos y deben ser descartados. Aunque en algunos campos y en ciertos casos este modo de actuar podría estar justificado, en muchas ocasiones resulta contraproducente.
En arqueología y otras áreas de la antropología, por ejemplo, hay desde hace tiempo encendidas disputas acerca de la difusión de los rasgos culturales frente a la invención independiente. Sin embargo, parece obvio que ocurren ambas cosas. La invención del cero en la India (desde donde llegó a Europa a través del trabajo de al Khwaritzmi) parece, con una evidencia abrumadora, haber sido independiente de su invención en Centroamérica (donde fue empleado por los antiguos mayas). Si hubieran existido contactos de alguna clase entre ambos mundos resultaría muy extraña la casi total ausencia de la rueda en las culturas precolombinas (por lo que sé sólo se ha encontrado en unos pocos juguetes de procedencia mejicana) cuando se conocía desde hace tanto tiempo en el Viejo Mundo. El arco y la flecha parecen haberse difundido de Norteamérica a Centroamérica, mientras que muchas otras innovaciones culturales, como la domesticación del maíz, lo hicieron en sentido contrario. ¿Cómo pueden los eruditos dividirse todavía en difusionistas y no difusionistas?
Algunos antropólogos culturales gustan de señalar las razones ecológicas y económicas que hay detrás de costumbres tribales que a primera vista pueden parecer arbitrarias o irracionales. Aunque este enfoque es muy valioso, a veces les lleva a ridiculizar la idea misma de que la irracionalidad y la arbitrariedad puedan jugar un importante papel en los sistemas de creencias y modelos de conducta social. Seguramente esto es ir demasiado lejos; un enfoque más razonable debería atemperar el determinismo ecológico y económico con una medida de la volubilidad de los esquemas tribales. Por ejemplo, una prohibición particular en la dieta, digamos la de comer okapis, podría tener sentido para cierta tribu, dadas las necesidades nutricionales de la población y el trabajo necesario para cazar un okapi en comparación con la obtención de otros alimentos en el contexto ecológico del bosque circundante. Pero la restricción también podría surgir de una identificación anterior del okapi con el tótem de la tribu; o bien ambas causas podrían estar en el origen de la prohibición. ¿Es acaso razonable insistir en que el enfoque correcto es siempre uno o es siempre el otro?
Una de las virtudes del Instituto de Santa Fe es que se ha creado un clima intelectual en el que eruditos y científicos se sienten mucho más inclinados que en sus instituciones de procedencia a compartir ideas y buscar maneras de armonizarlas en una síntesis fructífera cuando se considera conveniente. En una ocasión se organizó allí un seminario impartido por varios profesores de una misma universidad y un mismo departamento que comprobaron con cierto asombro que en Santa Fe se podía discutir constructivamente sobre temas que en su casa sólo provocarían disputas.
En la evolución biológica, lo más cercano a los esquemas cooperativos es probablemente la genética de la reproducción sexual, en la que los genotipos de los progenitores se mezclan en sus descendientes. Volveremos pronto a la reproducción sexual, pero primero exploraremos un poco más la tendencia hacia una mayor complejidad.
Hemos visto que la dinámica de la evolución biológica puede ser complicada. Pero a menudo ha sido descrita de manera simplista. La emergencia de formas cada vez más complejas se ha tomado más de una vez por una progresión continua hacia alguna clase de perfección, que suele identificarse con la especie, e incluso la raza o la estirpe, del autor de la idea. Por fortuna, este punto de vista teleológico está en decadencia, y hoy es posible contemplar la evolución más como un proceso que como un medio para alcanzar algún fin.
Aun así, todavía persiste, incluso entre algunos biólogos, la idea de que es inherente a la evolución biológica cierta «tendencia» hacia la complejidad. Como hemos visto, lo que realmente ocurre es algo más sutil. La evolución procede por pasos, y en cada paso la complejidad puede aumentar o disminuir, pero el efecto en la totalidad de especies existentes es que la mayor complejidad representada tiende a aumentar con el tiempo. Un proceso similar se da en las comunidades en crecimiento económico, donde una familia concreta puede ver cómo sus ingresos aumentan, disminuyen o no varían, pero la distribución de ingresos se va ampliando, de modo que los ingresos familiares máximos en el conjunto de la comunidad tienden a incrementarse.
Figura 18. Distribución cambiante de las distancias en un paseo aleatorio
Si ignorásemos cualquier ventaja atribuible a una complejidad incrementada, podríamos contemplar la distribución cambiante de complejidades como una especie de difusión, ejemplificada por un «paseo aleatorio» a lo largo de una línea. Un gran número de pulgas parten de un mismo punto y van dando saltos iguales en longitud, alejándose del punto de partida o acercándose a él al azar (en el primer salto, por supuesto, todas se alejan). En cualquier momento posterior, una o más pulgas estarán más lejos que las demás del punto de partida. Naturalmente, la pulga más distante en cada caso puede variar, dependiendo de cuál de ellas ha dado, por azar, el mayor número de saltos netos alejándose del punto de partida. La distancia máxima del conjunto de pulgas tiende a aumentar con el tiempo. La distribución de distancias desde el punto de partida se amplía a medida que la difusión de las pulgas las lleva cada vez más lejos, como se muestra en la Figura 18.
El crecimiento de la complejidad máxima puede proceder, especialmente en sistemas no adaptativos como las galaxias, de una manera que recuerda a la difusión. Pero en los sistemas complejos adaptativos, como la evolución biológica, suele ocurrir que las presiones selectivas favorecen una mayor complejidad en determinadas situaciones. La forma de la distribución de complejidades en función del tiempo diferirá en tales casos de la que resulta de un paseo aleatorio. Aunque sigue sin haber razón alguna para pensar en una progresión continua hacia organismos cada vez más complejos, las presiones selectivas que favorecen una complejidad más alta a menudo pueden ser intensas. La caracterización de aquellos sistemas y entornos físicos en los que la complejidad es una gran ventaja representa un desafío intelectual importante.
Los sucesos umbral que se presentan en el transcurso de la evolución biológica suelen dar lugar a grandes incrementos de complejidad y también a progresos muy significativos. La apertura de un umbral crítico se traduce en una explosión de nuevos nichos ecológicos, cuya ocupación puede muy bien parecer causada por una tendencia hacia una mayor complejidad.
Dado que nosotros los humanos somos los organismos más complejos en la historia de la Tierra, es comprensible que algunos contemplen la totalidad del proceso evolutivo como algo conducente al Homo sapiens sapiens. Aun reconociendo que esta idea responde únicamente a un necio antropocentrismo, en cierto sentido sí puede decirse que la evolución biológica termina con nosotros, o por lo menos queda en suspenso. Nuestro efecto sobre la biosfera es tan profundo y nuestra capacidad para transformar la vida (no sólo mediante procedimientos antiguos y lentos como los de los criadores de perros, sino con métodos modernos como la ingeniería genética) será pronto tan grande que ciertamente el futuro de la vida sobre la Tierra depende en gran parte de decisiones cruciales tomadas por nuestra propia especie. Salvo una espectacular renuncia a la tecnología (muy difícil a la vista de la enorme población humana que depende ya completamente de ella para su sustento) o la autodestrucción de la mayor parte del género humano —seguida de una regresión a la barbarie de los supervivientes— da la impresión de que, en un futuro previsible, el papel de la evolución biológica natural será, para bien o para mal, secundario al de la cultura humana y su evolución.
Desafortunadamente, pasará mucho tiempo antes de que el conocimiento, la inteligencia y el ingenio humanos puedan igualar —si es que lo hacen alguna vez— la «sabiduría» de miles de millones de años de evolución biológica. No sólo los organismos individuales han desarrollado morfologías intrincadas y modos de vida propios, sino que las interacciones entre la multiplicidad de especies que componen las comunidades ecológicas han experimentado delicados ajustes mutuos a lo largo de grandes periodos de tiempo.
Las diversas comunidades consisten en grupos de especies que difieren en función de la región del globo donde se encuentran y, dentro de cada región, del entorno físico. En tierra, el carácter de la comunidad varía de acuerdo con factores tales como la altitud, las lluvias y su distribución a lo largo del año, o la temperatura y sus variaciones. Las diferencias regionales se deben a que en muchos casos las distribuciones de las distintas especies se han visto afectadas por los movimientos de los continentes a lo largo de millones de años y accidentes tales como antiguas migraciones y dispersiones.
Los bosques tienden a diferir grandemente entre sí, incluso en los trópicos. No todos los bosques tropicales son pluvisilvas, como ciertos medios de comunicación podrían hacernos creer. Algunos son bosques secos, otros bosques de tipo alpino, etc. Por otra parte, pueden distinguirse cientos de pluvisilvas distintas, con diferencias significativas en la flora y en la fauna. Brasil, por ejemplo, alberga no sólo la vasta extensión de la selva amazónica, considerablemente variada en sí misma, sino también la muy diferente selva atlántica, ahora reducida a una pequeña fracción de su extensión inicial. En su margen meridional la selva atlántica se confunde con la del Alto Paraná en Paraguay y con la selva tropical de la provincia de Misiones en Argentina. La destrucción progresiva de la selva amazónica es ahora una preocupación general, aunque todavía se mantiene en pie gran parte de ella (a veces, por desgracia, en un estado degradado no detectable fácilmente desde el aire), pero la preservación de lo que queda de la selva atlántica es un asunto aún más urgente.
También los desiertos difieren entre sí. En el desierto del Namib, en Namibia, la flora y la fauna son en gran parte diferentes de las del desierto del Sahara, en el otro extremo de África, y de las del desierto espinoso del sur de Madagascar. Los desiertos de Mojave y del Colorado, en el sur de California, son bien distintos entre sí y comparten pocas especies con, digamos, el Negev israelí (una excepción notable es el sabra, el conocido cactus de Israel, que es una especie originaria de México y California). En una visita a los desiertos del Colorado y del Negev, una inspección superficial revelará muchas semejanzas en la apariencia de la flora, pero buena parte de este parecido es atribuible, no a un parentesco cercano entre las especies, sino a una convergencia evolutiva resultante de presiones selectivas similares. Igualmente, muchas euforbias de las mesetas áridas de África oriental se parecen a los cactus del Nuevo Mundo, pero sólo porque se han adaptado a climas similares; en realidad pertenecen a familias diferentes. La evolución ha producido en diversas partes del globo cierto número de soluciones distintas pero similares al problema representado por una comunidad de organismos viviendo en unas condiciones determinadas.
Ante tal diversidad de comunidades naturales ¿tendrán los seres humanos la sabiduría colectiva necesaria para elegir las políticas apropiadas? ¿Habremos adquirido el poder de efectuar cambios enormes antes de haber madurado lo suficiente como especie para hacer un uso responsable del mismo?
Las comunidades ecológicas compuestas de multitud de individuos complejos pertenecientes a gran número de especies, todos ellos desarrollando esquemas para describir y predecir el comportamiento mutuo, son sistemas que probablemente nunca alcanzan un estado estacionario final y ni siquiera se aproximan a él. Cada especie evoluciona en presencia de agrupaciones constantemente cambiantes de otras especies. La situación es muy diferente de la de los extremófilos oceánicos, que evolucionan en un entorno fisicoquímico prácticamente constante e interaccionan con otros organismos principalmente a través de la materia orgánica que desciende hasta ellos a través del agua.
Ni siquiera a un sistema comparativamente simple y casi autocontenido como el de los extremófilos se le puede asignar un atributo numérico rigurosamente definido que se llame «adaptación», y mucho menos que vaya aumentando en el curso de la evolución hasta llegar a un estado estacionario. Incluso en un caso tan simple es más seguro concentrarse directamente en las presiones selectivas que favorecen unos caracteres fenotípicos sobre otros, influyendo así en la competencia entre los diferentes genotipos. Estas presiones selectivas pueden no ser expresables en términos de una magnitud simple y bien definida llamada adaptación, sino que pueden requerir una descripción más complicada, incluso en el caso ideal de una sola especie que se adapta a un medio ambiente inmutable. Todavía es menos probable, pues, que pueda asignarse una medida de adaptación a un organismo en un entorno cambiante, especialmente cuando se trata de un miembro de una comunidad ecológica de organismos altamente interactivos adaptándose a las peculiaridades de unos y otros.
Aún así, un tratamiento simplificado de la evolución biológica en términos de adaptación puede resultar instructivo. La idea subyacente al concepto de adaptación biológica es que la propagación de los genes de una generación a la siguiente depende de la supervivencia del organismo hasta que éste alcanza la fase reproductora y genera un número razonable de descendientes que a su vez sobreviven para reproducirse. Las tasas diferenciales de supervivencia y reproducción pueden describirse a menudo toscamente en términos de un índice de adaptación, definido de manera que los organismos con una mayor adaptación tienden en general a propagar sus genes con más éxito que los que tienen una adaptación menor. En el límite, los organismos con patrones genéticos asociados consistentemente con el fracaso reproductivo tienen una adaptación muy baja y tienden a desaparecer.
Al introducir la tosca noción de «relieve adaptativo» se pone de manifiesto una dificultad general. Imaginemos los diferentes genotipos dispuestos en una superficie bidimensional horizontal (que representa lo que en realidad es un espacio matemático multidimensional de genotipos posibles). La adaptación o inadaptación queda indicada por la altura; a medida que los genotipos varían la adaptación describe una superficie bidimensional, con gran número de crestas y valles, dentro de un espacio de tres dimensiones. Los biólogos representan convencionalmente el incremento de la adaptación como un incremento de altura, de modo que los máximos de adaptación corresponden a las cimas y los mínimos al fondo de los valles; pero yo emplearé la convención inversa, como se acostumbra a hacer en muchos otros campos, y daré la vuelta a todo el cuadro. Ahora la adaptación aumenta con la profundidad, y los máximos se encuentran en el fondo de las depresiones, como se muestra en la Figura 19.
El paisaje que resulta es muy complicado, con numerosos valles («máximos de adaptación locales») de profundidad muy variable. Si la evolución tuviese siempre como efecto un descenso continuo —una mejora constante de la adaptación— entonces el genotipo quedaría probablemente atrapado en el fondo de una depresión somera y no tendría oportunidad de alcanzar los valles profundos cercanos que corresponden a una adaptación mucho mayor. El movimiento del genotipo tiene que ser, como mínimo, más complicado que un simple deslizamiento hacia abajo. Si además incluyese, por ejemplo, saltos aleatorios, esto le daría la oportunidad de escapar de las depresiones someras y encontrar otras más profundas en las cercanías. Sin embargo, no debe haber demasiados saltos, o todo el proceso dejaría de funcionar. Como hemos visto en diversos contextos, un sistema complejo adaptativo funciona mejor en una situación intermedia entre el orden y el desorden.
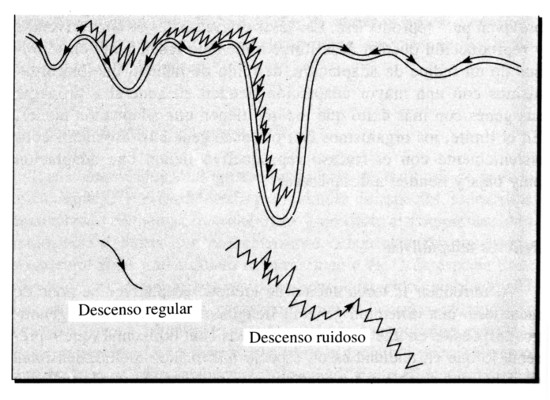
Figura 19. Un relieve adaptativo en el que la adaptación crece con la profundidad
Una complicación adicional en el manejo del concepto de adaptación surge en los organismos superiores que se reproducen sexualmente. Cada uno de tales organismos transfiere sólo la mitad de sus genes a la descendencia, mientras que la otra mitad procede del otro progenitor. Los descendientes no son clones, sino meros parientes cercanos. Y el organismo considerado tiene también parientes cercanos, la supervivencia de los cuales puede contribuir de alguna manera a la propagación de genes similares a los suyos. Para tener en cuenta hasta qué punto sobreviven y consiguen reproducirse los parientes de un organismo dado, de acuerdo con la cercanía del parentesco, los biólogos han desarrollado la noción de «adaptación inclusiva» (naturalmente, la adaptación inclusiva también tiene en cuenta la supervivencia del organismo mismo). La evolución debería mostrar una tendencia general a favorecer los genotipos que exhiban una alta adaptación inclusiva, especialmente a través de pautas de comportamiento heredadas que promuevan la supervivencia tanto del organismo individual como de sus parientes cercanos. Esta tendencia recibe el nombre de «selección de parentesco», y encaja bien en un cuadro evolutivo en el que los organismos serían meros instrumentos de los que los genes «se sirven» para propagarse. Este punto de vista ha sido popularizado como el del «gen egoísta».
Una forma extrema de gen egoísta podría ser responsable del fenómeno conocido como «distorsión segregacional». Descrita por el sociobiólogo Robert Trivers, la distorsión segregacional resultaría de la operación de un «gen literalmente egoísta», es decir, un gen que actúa directamente, no a través del organismo resultante, para triunfar en la competencia con los patrones genéticos rivales. Uno de tales genes presente en un animal macho podría hacer que sus espermatozoides adelantasen o incluso envenenasen a los de otros machos, haciendo así más fácil la victoria en la carrera para fecundar los huevos de la hembra. En cambio, un gen literalmente egoísta no tiene por qué conferir ninguna ventaja al organismo resultante, e incluso podría ser nocivo en cierto grado.
Aparte de estas notables excepciones, las presiones selectivas se ejercen indirectamente a través del organismo producido por la fusión de espermatozoide y óvulo. Esto está más en línea con la noción de sistema complejo adaptativo, donde el esquema (en este caso el genoma) se pone a prueba en el mundo real (por medio del fenotipo) y no directamente.
Un caso fascinante en el que parecen estar implicadas tanto la adaptación individual como la inclusiva es el denominado comportamiento altruista en ciertas especies de aves. El arrendajo mejicano vive en los hábitats áridos del norte de México, sureste de Arizona y suroeste de Nuevo México. Hace años los ornitólogos observaron que cada nido de esta especie era atendido a menudo por varias aves además de la pareja que produjo los huevos. ¿Qué hacían allá los otros arrendajos? El comportamiento que exhibían ¿era en verdad altruista? La investigación de Jerram Brown reveló que en muchos casos los ayudantes eran ellos mismos hijos de la pareja que criaba, y por lo tanto estaban ayudando a criar a sus propios hermanos. Este comportamiento parecía suministrar un notable ejemplo de evolución del comportamiento social a través de la adaptación inclusiva. La evolución había favorecido un comportamiento paternal en el que los arrendajos jóvenes, posponiendo su propia reproducción, ayudaban a alimentar y cuidar a sus hermanos menores, asistiendo así a la propagación de genes estrechamente relacionados con los propios.
Más recientemente, el cuadro se ha complicado algo a raíz de los trabajos de John Fitzpatrick y Glen Woolfenden acerca de una especie emparentada, el arrendajo de Florida, que habita en los robledales áridos —en franca recesión— del sur de aquella región. Hasta ahora esta ave se consideraba una de las muchas subespecies del arrendajo azul americano, muy común en el sudoeste de los Estados Unidos, pero Fitzpatrick y Woolfenden proponen que debe considerarse una especie separada, atendiendo a su apariencia, vocalizaciones, comportamiento y genética. Como en el arrendajo mejicano, el comportamiento de esta especie incluye la crianza asistida. Aquí también los ayudantes tienden a ser hijos de la pareja que cría, pero las observaciones de los investigadores de Florida indican que los ayudantes actúan también en interés propio. Los territorios de anidamiento en el robledal árido son extensos (del orden de treinta acres) y son defendidos con fiereza, por lo que no resultan fáciles de conseguir. Los ayudantes están en la mejor posición para heredar todo o parte del territorio donde desempeñan su labor. Por lo menos en Florida, parece que la adaptación individual ordinaria tiene un papel muy importante en el comportamiento «altruista» del arrendajo.
No he presentado la historia del arrendajo de Florida para tomar partido en una controversia entre ornitólogos, sino para ilustrar la sutileza del concepto de adaptación, inclusiva o no. Aunque es un concepto útil, sigue siendo un tanto circular. La evolución favorece la supervivencia de los mejor adaptados, y los mejor adaptados son los que sobreviven, o aquellos cuyos parientes próximos sobreviven.
El fenómeno de la reproducción sexual plantea algunos retos especiales a las teorías basadas en las presiones selectivas y la adaptación. Como muchos otros organismos, los animales superiores tienden a reproducirse sexualmente. Pero en ciertos casos esos mismos animales pueden reproducirse facultativamente por partenogénesis, un proceso en el que las hembras, sin requerir los servicios de un macho, generan hembras hijas que, aparte de posibles mutaciones, son genéticamente idénticas a las progenitoras. Incluso los huevos de un animal tan complejo como la rana pueden producir, tras ser estimulados con la punta de una aguja, renacuajos que se desarrollan hasta convertirse en ranas adultas. En algunos casos raros, como el de los lagartos de cola de látigo de México y el sudoeste de los Estados Unidos, una especie de vertebrado parece arreglárselas sólo con la partenogénesis, sin que haya machos en absoluto. ¿Para qué el sexo entonces? ¿Qué enorme ventaja confiere la reproducción sexual? ¿Por qué se selecciona en general frente a la partenogénesis? ¿Para qué sirven en verdad los machos?
La reproducción sexual introduce diversidad en los genotipos de los descendientes. A grandes rasgos, los cromosomas (cada uno con una cadena de genes) se presentan en pares, y cada individuo hereda uno de los componentes de cada par del padre y el otro de la madre. Cuál de ellos procede del padre y cuál de la madre es fundamentalmente una cuestión de azar. (En los gemelos idénticos, el resultado de esta repartición estocástica es el mismo para ambos.) La descendencia de los organismos con muchos pares de cromosomas tiene por lo general juegos de cromosomas diferentes de los de los progenitores.
Además de eso, la reproducción sexual introduce un mecanismo completamente nuevo, distinto de la mutación ordinaria, para producir cambios en los cromosomas. En el proceso denominado «entrecruzamiento», ilustrado en la Figura 20, un par de cromosomas homólogos intercambian fragmentos durante la formación de un espermatozoide o un huevo. Supongamos que el entrecruzamiento tiene lugar en el segundo caso. El huevo, producido por la madre, adquiere un cromosoma mixto, una parte del cual es aportada por el padre de la madre y el resto por la madre de la madre, mientras que otro huevo puede recibir un cromosoma formado por las partes restantes de los cromosomas del abuelo y la abuela matemos.
El teórico evolucionista William Hamilton, ahora catedrático en Oxford, ha sugerido una explicación simple para el valor de la reproducción sexual. En términos generales, la idea es que para los enemigos de una especie, especialmente los parásitos nocivos, resulta más difícil adaptarse a la diversidad de una población generada por reproducción sexual que a la relativa uniformidad de una población generada por partenogénesis. La mezcla de los cromosomas aportados por el padre y la madre, junto con el entrecruzamiento, permiten todo tipo de combinaciones nuevas entre los descendientes, obligando a los parásitos a enfrentarse a una gran variedad de huéspedes diferentes en su química corporal, hábitos, etc. El resultado es que los enemigos encuentran dificultades y los huéspedes están más seguros.
La teoría indica que las especies sin reproducción sexual deberían disponer de otros mecanismos para enfrentarse a los parásitos, especialmente en el caso de grupos enteros de animales inferiores que han vivido sin sexo durante decenas de millones de años. Los rotíferos bdeloideos son uno de tales grupos. Son animalillos que se desplazan girando y habitan en las alfombras de musgo y otros lugares que permanecen húmedos la mayor parte del tiempo, pero que se secan durante semanas o meses según los caprichos de la meteorología. Una discípula de Hamilton, Olivia Judson, está estudiando estos rotíferos para averiguar cómo se las arreglan con los parásitos. Ella sugiere que su hábito de desecarse y dejarse llevar por el viento cuando su entorno se seca podría proporcionarles la suficiente protección contra los parásitos como para prescindir del sexo.
En cualquier caso, las ventajas de la reproducción sexual tienen que ser considerables para compensar la desventaja obvia que supone la fragmentación de los genotipos de padres y abuelos con una capacidad demostrada para sobrevivir y reproducirse con éxito. Estas ventajas son relativas al conjunto de la población, aunque muchos evolucionistas insisten en que las presiones selectivas se ejercen únicamente sobre los individuos. Pero quizás ésta no sea una norma rígida.
En un reciente encuentro en el Instituto de Santa Fe, John Maynard Smith, que enseña en la Universidad de Sussex, comentaba este tema cuando Brian Arthur, presidente de la sesión, recordó la ocasión en que ambos se encontraron por primera vez. Los dos estudiaron la carrera de ingeniería. Maynard Smith comenzó diseñando aviones y después se pasó a la biología evolutiva, donde ha hecho importantes contribuciones. Brian, que se crió en Belfast, se dedicó a la investigación operativa y más tarde a la economía. Después se convertiría en profesor en Stanford y sería el director fundador del programa económico del Instituto de Santa Fe. Se encontraron por primera vez en un congreso científico en Suecia, donde en el curso de su intervención Maynard Smith hizo notar que mientras el sexo tenía ventajas obvias para las poblaciones, no estaba claro que las tuviera para los individuos. Brian gritó desde la audiencia: «¡Vaya visión del sexo más inglesa!». Maynard Smith, tras encajar el golpe, replicó: «Deduzco por su acento que usted es irlandés. Bien, en Inglaterra por lo menos tenemos sexo».
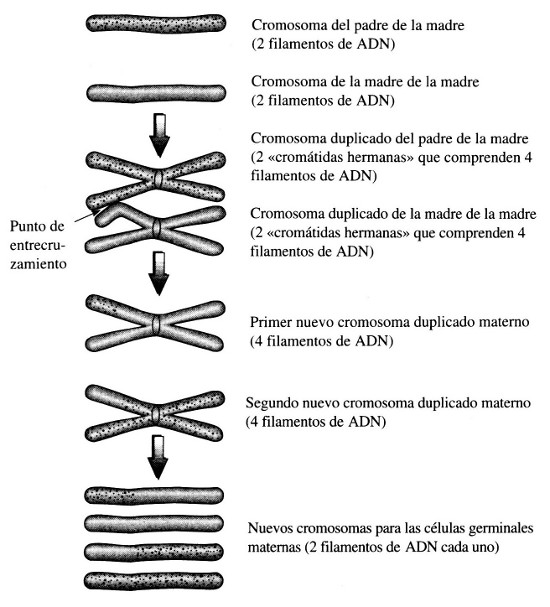
Figura 20. Entrecruzamiento de cromosomas en la reproducción sexual
Mientras que el sexo no es algo ni mucho menos universal en biología, la muerte sí que está muy cerca de serlo. La muerte de los organismos es una de las más dramáticas manifestaciones de la segunda ley de la termodinámica. Y como tal es, en cierto sentido, común a todos los sistemas complejos adaptativos. Sin embargo, es especialmente significativa en la evolución biológica, donde la interacción entre muerte y reproducción está en la vanguardia del proceso adaptativo. La competencia entre grupos de genotipos se traduce en gran medida en la competencia por el tamaño de la población entre los tipos de organismos correspondientes. La adaptación, hasta donde puede hacerse una buena definición de ella en el contexto de la evolución biológica, está siempre conectada con el tamaño de la población.
La comparación entre diversas clases de sistema complejo adaptativo revela situaciones en que la muerte, la reproducción y la población tienen menos importancia que en biología. Por ejemplo, imaginemos un ser humano individual absorto en la resolución de un problema. En este caso los esquemas son ideas y no genotipos. El análogo de la muerte es el olvido. Nadie puede negar la omnipresencia y significación del olvido, pero difícilmente puede compararse su papel con el de la muerte en biología. Si no fuese necesario olvidar, «borrar la cinta», el carácter del pensamiento no experimentaría grandes cambios. El recuerdo de una idea es útil, y contrarresta el efecto del olvido, pero el número de recuerdos idénticos o casi idénticos no caracteriza la adecuación de una idea en la misma medida que la población tiende a correlacionarse con la adaptación en biología.
A medida que las ideas se propagan en el seno de una sociedad (también en la comunidad científica) adquiere importancia el número de personas que comparten una idea determinada. En las elecciones democráticas, en la medida en que tienen que ver con las ideas, la opinión mayoritaria es la que prevalece. Pero está claro que la existencia de un abrumador número de partidarios no necesariamente implica que una idea sea correcta, y ni siquiera garantiza su supervivencia a largo plazo.
Un ejemplo más cercano a la evolución biológica puede ser el de la competencia entre las sociedades humanas en el pasado. En gran medida, la adaptación se medía por la población. En el Sudeste Asiático, por ejemplo, algunos grupos étnicos practicaban la agricultura de regadío para el cultivo del arroz, mientras que otros practicaban una agricultura de secano, a menudo quemando el bosque. Los primeros, como los pueblos de Tailandia central, Laos o Vietnam, podían mantener muchos más individuos por unidad de superficie que sus vecinos. La mayor densidad de población les permitió dominar a los agricultores de secano, que en muchos casos se vieron relegados a tierras montañosas remotas. De cara al futuro, bien podemos preguntarnos si es deseable que la densidad o la población absoluta continúen determinando de la misma manera quiénes ganan y quiénes pierden.
A largo plazo, la evolución biológica, con su énfasis en la muerte y la población, es perfectamente eficiente en la ocupación de nuevos nichos ecológicos a medida que éstos surgen. Cuando existe la oportunidad de adoptar un cierto modo de vida, es probable que algún organismo evolucione para aprovecharla, por estrafalario que pueda parecer a los ojos de un observador humano.
En este contexto es relevante la analogía entre una comunidad ecológica y la economía de mercado. Cuando surgen nuevas oportunidades para hacer negocio en una economía de este tipo, lo más probable (aunque no es seguro) es que aparezcan individuos o empresas dispuestos a explotarlas. La analogía de la muerte en este caso es la quiebra, y es la riqueza, en vez de la población, lo que da una medida aproximada de la adaptación de la empresa.
Tanto en economía como en ecología, la aparición de un nuevo negocio o un nuevo organismo (o de una nueva forma de actuar en una empresa u organismo ya existente) altera el relieve adaptativo de los otros miembros de la comunidad. Desde el punto de vista del negocio o de la especie, este relieve experimenta continuos cambios (además de no estar del todo bien definido a priori).
Ambos casos ilustran cómo un sistema complejo adaptativo, una vez establecido, puede llenar nichos, crear otros nuevos en el proceso, llenar éstos a su vez y así sucesivamente, engendrando nuevos sistemas complejos adaptativos por el camino. (Como se indicaba en el diagrama de la Figura 1, la evolución biológica ha dado lugar al sistema inmunitario de los mamíferos, al aprendizaje y al pensamiento y, a través de los seres humanos, a sociedades capaces de aprender y adaptarse, y más recientemente a ordenadores que funcionan como sistemas complejos adaptativos.)
Siempre explorando, buscando nuevas oportunidades, experimentando con la novedad, el sistema complejo adaptativo ensaya incrementos de complejidad y ocasionalmente descubre sucesos umbral que abren posibilidades estructurales completamente nuevas, incluyendo nuevas formas de sistema complejo adaptativo. Dado un tiempo suficiente, la probabilidad de la evolución de la inteligencia parece que debiera ser alta.
Astrónomos y planetólogos piensan que no existe ninguna razón para pensar que los sistemas planetarios sean especialmente raros en nuestra galaxia o en cualquier otra galaxia similar en el universo. Tampoco los teóricos del origen de la vida tienen motivos para pensar que las condiciones de nuestro planeta hace unos 4000 millones de años fueran tan especiales que la aparición de la vida (o algo parecido a ella) en un planeta sea un acontecimiento particularmente improbable. Es verosímil que en el universo abunden los sistemas complejos adaptativos y que en muchos de ellos haya evolucionado, o acabe por evolucionar, la inteligencia. Como ya hemos mencionado, para la búsqueda de inteligencia extraterrestre el principal dato que falta es el número de planetas por unidad de volumen espacial en los que han surgido seres inteligentes y la duración típica del período de civilización técnica con emisión de señales electromagnéticas. Dada la inmensa cantidad de conocimiento que podemos extraer de la diversidad de comunidades naturales en la Tierra, por no hablar de la diversidad de sociedades humanas, resulta sobrecogedor imaginar (como hacen a veces los escritores de ciencia ficción) las lecciones que el contacto con extraterrestres podría damos acerca de la variedad de circunstancias que los sistemas complejos adaptativos son capaces de explotar.
Para encontrar ejemplos curiosos de explotación de oportunidades por parte de especies en interacción con otras, podemos fijamos en el engaño tal como es practicado por animales no humanos. El mimetismo es bien conocido; la mariposa virrey, por ejemplo, se parece a la monarca y se aprovecha así del rechazo que inspira ésta en los depredadores a causa de su mal sabor. El cuclillo (en el Viejo Mundo) y el boyero (en el Nuevo Mundo) practican otro tipo de engaño, poniendo sus huevos en los nidos de otras aves; los polluelos intrusos se deshacen después de los huevos o polluelos legítimos y monopolizan la atención de los padres adoptivos. Ahora bien, ¿realmente engañan?
Estamos acostumbrados a oír que la gente engaña, pero que esto ocurra en otros organismos resulta más sorprendente. Cuando la marina argentina descubre un misterioso periscopio en el estuario del Río de la Plata justo antes de que el presupuesto de las fuerzas armadas comience a debatirse, podemos sospechar que se está intentando engañar para conseguir recursos adicionales, lo que no nos resulta especialmente sorprendente. Pero la existencia de un comportamiento análogo entre las aves es algo inesperado.
Un caso así fue descubierto recientemente por mi amigo Charles Munn, un ornitólogo que estudia las bandadas mixtas que se alimentan en las selvas bajas tropicales del Parque Nacional Manu, en Perú. Algunas especies ramonean juntas en el sotobosque y otras en el estrato inferior del dosel, acompañadas de vez en cuando por coloreados tangarás frugívoros procedentes del estrato superior. (Entre las especies presentes en estas bandadas en la época invernal hay unos pocos migradores norteamericanos. Más al norte, hacia Centroamérica, se encuentran muchos más. Quienes residimos en Norteamérica sólo las vemos cuando anidan en verano, y nos llama la atención encontrarlas llevando un modo de vida muy diferente en tierras lejanas. Si queremos que vuelvan para anidar año tras año, hay que proteger sus hábitats en los países meridionales. Igualmente, su retorno a estos países quedará comprometido si los bosques norteamericanos continúan siendo reducidos a parcelas cada vez más pequeñas. La reducción de los bosques propicia invasiones posteriores por parte de los boyeros parásitos.)
En cada bandada mixta hay una o dos especies centinela que deambulan de manera que suelen estar en medio de la bandada o justo debajo. Mediante un canto especial los centinelas advierten a los demás de la presencia en las proximidades de posibles aves rapaces. Charlie observó que los centinelas de las bandadas del sotobosque daban a veces la señal de aviso aun cuando no existía un peligro aparente. Una inspección más cercana demostraba que la falsa alarma permitía a menudo al centinela arrebatar un suculento bocado que de otro modo se habría comido otro miembro de la bandada. Una observación cuidadosa reveló que los centinelas practicaban el engaño alrededor de un 15% de las veces, sacando provecho de él con frecuencia. Preguntándose si este fenómeno sería más general, Charlie examinó el comportamiento de las bandadas del estrato inferior del dosel y encontró que los centinelas de allá hacían lo mismo. El porcentaje de señales falsas era más o menos el mismo en las dos especies centinela. Presumiblemente, si este porcentaje fuera mucho mayor las señales dejarían de ser aceptadas por el resto de la bandada (recuérdese el cuento del pastorcillo que gritaba «¡que viene el lobo!») y si fuera mucho menor, el centinela apenas obtendría alimento extra. Me pregunto si este dato del 15% podría derivarse de alguna clase de razonamiento matemático; en un modelo plausible, ¿podría corresponder a 1/2π?
Cuando consulté esta cuestión con Charles Bennett, recordó algo que le había contado su padre sobre las unidades de la fuerza aérea canadiense con base en Inglaterra durante la segunda guerra mundial. Para las ocasiones en que enviaban un caza y un bombardero juntos, les pareció una buena idea intentar engañar de vez en cuando a la Luftwaffe haciendo volar el caza por debajo del bombardero y no por encima como era lo habitual. Tras aplicar el método de ensayo y error un buen número de veces, llegaron a la conclusión de que lo mejor era hacerlo al azar una vez de cada siete.
En nuestra discusión de los sucesos umbral citábamos algunos ejemplos de avances en la evolución biológica que representaban saltos enormes, pero también indicábamos que se trata de acontecimientos raros en el extremo de todo un espectro de cambios de magnitud diversa, siendo mucho más comunes los cambios pequeños cerca del otro extremo del espectro. Sea cual sea la magnitud del evento, la evolución biológica suele proceder actuando sobre lo que hay disponible. Los órganos existentes se adaptan a nuevos usos. Los brazos humanos, por ejemplo, no son más que patas anteriores ligeramente modificadas. Las estructuras no son radicalmente descartadas en un rediseño revolucionario de la totalidad del organismo. Los mecanismos de la mutación y la selección natural no favorecen tales discontinuidades. Pero las revoluciones ocurren.
Hemos discutido cómo, en el fenómeno del «equilibrio puntuado», los cambios relativamente repentinos pueden tener orígenes diversos. Uno de los posibles es un cambio en el entorno fisicoquímico que altere significativamente las presiones selectivas. Otro es la «deriva», en la que las mutaciones neutras, que no alteran la viabilidad del fenotipo (y a veces ni siquiera éste) conducen gradualmente a una situación de inestabilidad del genotipo, donde una o unas pocas mutaciones pueden crear un organismo significativamente diferente y preparar el camino para una cascada de cambios que se extiendan a otras especies. A veces pequeños cambios desencadenan sucesos umbral, muchas veces de carácter bioquímico, que abren dominios enteros de formas de vida completamente nuevas. En algunos casos estos cambios revolucionarios se inician con la agregación de organismos en estructuras compuestas. Pero en todos los casos la unidad básica de cambio es una mutación (o recombinación, con o sin entrecruzamiento) que opera sobre lo que ya está presente. Nunca se saca nada de donde no hay.
¿Hasta qué punto es éste un principio general para los sistemas complejos adaptativos? En el pensamiento humano, por ejemplo, ¿es necesario proceder por pequeños pasos? ¿Debe limitarse el proceso de creación a encadenamientos de cambios menores sobre lo ya existente? ¿Por qué no podría un ser humano ser capaz de inventar un ingenio totalmente nuevo, diferente de cualquier otro conocido? Y en ciencia, ¿por qué no concebir una teoría completamente nueva, sin ninguna semejanza con las ideas precedentes?
La investigación (y también la experiencia diaria) parece indicar que, efectivamente, el pensamiento humano suele proceder por asociación y por pasos, en cada uno de los cuales se introducen modificaciones específicas sobre lo pensado con anterioridad. Pero tanto en la invención como en la ciencia, el arte, y muchos otros campos de la creación humana, de vez en cuando surgen estructuras singularmente novedosas. Tales innovaciones recuerdan los sucesos umbral en la evolución biológica. ¿Cómo se producen? ¿Responde el pensamiento creativo humano a modelos diferentes según las diferentes áreas de actividad? ¿O hay involucrados principios generales de alguna clase?