Aunque Sydney y yo nos dábamos cuenta de que el código genético era un problema bioquímico, todavía teníamos esperanzas de que los métodos genéticos pudieran contribuir a la solución, ya que si se utiliza el material correcto los métodos genéticos suelen ser rápidos, mientras que los métodos bioquímicos son a menudo bastante lentos. Seymour Benzer había utilizado métodos genéticos para demostrar que el material genético era seguramente unidimensional. La cuestión había sido inspirada por la doble hélice del DNA, pero el método utilizado era enteramente original.
Cuando se quiere mapear un gen en profundidad es necesario tomar individuos bastante extraños. Cuanto más cerca se encuentren dos mutantes en un gen, más raro será el hecho de la recombinación genética entre ellos. Benzer había escogido un sistema con dos ventajas. Los genes en cuestión se encontraban en el bacteriófago T4, un virus que atacaba y mataba las células de E. coli. El virus crece rápido y recombina a alta velocidad. Había escogido un gen llamado rII (de hecho se trataba de un par de genes cercanos) porque presentaba una ventaja técnica muy especial. Utilizando estirpes apropiadas de la célula huésped era posible detectar un virus que tuviera un gen silvestre incluso si estaba mezclado con millones de virus con la versión mutante. De esta forma, podían ser detectados ciertos recombinantes muy raros, tan raros que Benzer calculó que se podrían separar incluso pares de bases adyacentes sobre el DNA. Por desgracia, no había un método semejante para encontrar un mutante entre un gran número de tipos silvestres, pero con el huésped apropiado, la calva (la pequeña colonia formada por el crecimiento de una única bacteria con virus sobre una cepa de E. coli en una placa de Petri) era muy diferente y fácil de reconocer. Un único mutante en una placa de Petri conteniendo varios centenares de calvas silvestres era susceptible de detectarse de manera bastante fácil.
Se podría haber seguido el método usual, tomando una serie de mutantes distintos para encontrar la distancia de recombinación entre cualquier par de ellos. Eran también posibles métodos más elaborados utilizando tres mutantes, pero todos ellos implicaban el recuento de centenares o millares de calvas, lo que era muy laborioso.
Benzer, quien siempre intenta ahorrarse el trabajo innecesario, pensó en un método mejor. Además de mutaciones puntuales encontró que algunos de sus mutantes parecían ser deleciones. Estas mapeaban como trazos en su mapa genético, ya que parecían solapar con al menos dos mutaciones puntuales. De esta forma podía recoger varias series completas de deleciones. Si dos deleciones solapaban, la recombinación genética nunca podría volver a dar el tipo silvestre intacto ya que la porción solapante no estaba en ninguno de los padres y por ende no podían ser recuperadas. Por otra parte, si las dos deleciones no solapaban, una recombinación apropiada podría dar el tipo silvestre.
Una analogía ayudará a aclarar lo antedicho. Imaginemos dos copias defectuosas de un libro, uno al que le falta de la página 100 a la 120 y el otro al que le falta de la 200 a la 215. Evidentemente, con estas dos copias, cada una con una deleción contigua única, podríamos recuperar el texto del libro por completo. Sin embargo, si al segundo libro, en lugar de faltarle las páginas 200 a 215 le faltaran las páginas 110 a 125, no habría manera de recuperar las páginas 110 a 120, ya que éstas faltan en las dos copias.
Para que la analogía sea aún más aproximada tenemos que ampliarla un poco. Imaginemos que el libro contenía instrucciones muy detalladas para hacer un instrumento complicado. Supongamos también que con cualquier página que faltara no podría hacerse el instrumento y si se hiciera, no funcionaría. Finalmente supongamos que tuviéramos millones de copias de cada uno de los libros defectuosos. La regla sería entonces: seleccionemos un ejemplar de cada tipo de libros. A continuación las primeras n páginas de un libro y las restantes páginas que quedan del otro. Miremos si este nuevo libro híbrido produce un instrumento que funcione. Hagamos esto millones de veces, y seleccionemos cada vez n páginas al azar. Si ocasionalmente se produce un buen instrumento, las dos deleciones no solapan. Si no se produce nunca un buen instrumento, entonces probablemente las dos deleciones se solapan.
Quizás ésta parezca una manera demasiado elaborada de hacerlo, pero ya que no podíamos mirar dentro del fago, era la única que teníamos. Todo lo que Benzer tenía que hacer entonces era juntar dos virus infectando simultáneamente un cultivo de E. coli con ellos. Después de que hubieran crecido y recombinado dentro de la bacteria, los virus podían ser plaqueados sobre una placa de Petri con una estirpe apropiada. Si las deleciones no solapaban, entonces habría algunas calvas recombinantes sobre la placa de Petri. Si solapaban, no habría ninguna. No había necesidad de un recuento laborioso, todo lo que se necesitaba era una respuesta de sí o no.
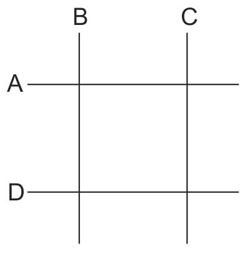
El razonamiento de Benzer era que si un gen tenía dos dimensiones, debíamos encontrar un patrón especial cuando hay cuatro deleciones. La deleción A se solaparía con la B y la C y también la D, pero las deleciones B y C no podrían solaparse ni tampoco podrían hacerlo la A y la D. (Véase la figura 12.1.) Es fácil percibir que esto no podría ocurrir si el gen era de una sola dimensión. Benzer tomó centenares de deleciones y las cruzó una con otra en pares. La situación que se presenta en la figura nunca ocurrió. Por tanto, concluyó, el gen es probablemente unidimensional. Sus resultados también le permitieron poner todas sus deleciones en orden y de esta forma se podía ver aproximadamente dónde mapeaba cada una de ellas en el gen.
Figura 12.1. Demostración de que en dos dimensiones podemos tener A solapada con B y C a la vez, y D también solapada con B y C, sin que A solape a D o B solape a C. Esto sería imposible si A, B, C y D fueran segmentos de una línea recta (unidimensional). Dado que Benzer no encontró este patrón de solapamiento en su colección de deleciones, llegó acertadamente a la conclusión de que el gen que estaba estudiando era unidimensional, hecho compatible con que estuviera constituido por DNA.
Por una serie de razones, nuestro grupo había escogido el mismo sistema de trabajo. Nuestro principal interés radicaba en los diferentes tipos de mutantes producidos por diferentes sustancias químicas y también en las reversiones que estas sustancias químicas producían. Los mutantes parecían repartirse en dos clases. Muchas sustancias químicas producían mutantes de la primera clase. Sin embargo los mutantes producidos por las sustancias químicas del tipo acridina eran de la otra. Cada clase se eliminaba más fácilmente usando el mismo tipo de mutágeno que la había producido. Ernst Freese había sugerido que una clase correspondía a transiciones (purina a purina o pirimidina a pirimidina, véase apéndice A) mientras que la otra correspondía a transversiones (purina a pirimidina o viceversa). Nosotros estábamos trabajando con otra idea. Algunos mutantes tenían filtraciones (es decir, el gen estaba activo de alguna manera aunque no totalmente activo), mientras que otros no tenían filtraciones, es decir, no tenían esencialmente ninguna actividad. Nos dimos cuenta de que los mutantes producidos por la proflavina (una acridina típica) eran casi siempre sin filtraciones. Esto nos llevó a sugerir que los mutantes de proflavina eran pequeñas deleciones o adiciones a la secuencia de bases, mientras que todas las otras clases de mutantes eran sustituciones de bases de un tipo o del otro. Sin embargo, aún nos faltaba una evidencia más clara para confirmar esta idea.
Mientras tanto yo había llegado a una idea muy diferente. Pensando en cómo una molécula de RNA puede trabajar como mensajera, dudaba si podía plegarse sobre sí misma formando una estructura en doble hélice más o menos débil. Esta idea consistía en que algunas bases podían aparearse mientras las otras, las que no se unen según la regla del apareamiento, formarían lazos hacia fuera. El «código» dependería entonces de si las bases formaban lazos o pares, o quizás alguna combinación más elaborada de estas dos posibilidades. Esta idea era en realidad muy vaga, pero permitía hacer una predicción muy importante. Al recibir el mensaje, un mutante podría en teoría ser capaz de ser compensado en su defecto por otro situado hacia el otro extremo con el cual se apareaba. Así, algunos mutantes podrían tener los llamados «supresores» distantes dentro de un mismo gen.
Esta idea me gustaba bastante aunque nadie se ocupó mucho de ella. Por aquel tiempo ya no hacía genética de fagos, simplemente me contentaba con observar los resultados de mis colegas. Ya que nadie parecía considerar la posibilidad de probar mi idea, me decidí a probarla yo mismo. No fue muy difícil aprender genética de fagos, especialmente al lado de una mano experta. A pesar de ello, cometí algunos errores elementales que, afortunadamente, corregí muy pronto. Los experimentos también me enseñaron cuán superficial era mi conocimiento, incluso a pesar de haber participado en numerosas discusiones sobre este mismo sistema. No hay nada mejor que la experimentación activa de uno mismo para darse cuenta de los entresijos de una técnica. Esto ayuda también a fijar los detalles en la propia mente, sobre todo porque leer la sección de «métodos experimentales» de la mayoría de los artículos científicos es la cosa más aburrida que puede imaginarse.
Naturalmente escogí los genes rII para el experimento concentrándome en el segundo, el llamado cistrón B. (Cistrón era el nombre que Benzer había encontrado para un gen definido gracias al test denominado cis-trans). Seleccioné un mutante de nuestro stock, intenté encontrar un revertiente (uno que se parecía al tipo silvestre) y después miré si esta reversión era debida a una segunda mutación en alguna parte del mismo gen. Si no la encontraba, continuaba y probaba otra vez con otro mutante.
Al principio no logré encontrar ningún supresor. Era posible que el cambio que revertía el mutante al tipo silvestre estuviera muy cerca del lugar original o demasiado cerca para que yo pudiera observarlo. Un día llegó Leslie Orgel a la hora del café. Mientras él miraba por encima de mi hombro le expliqué lo que estaba haciendo y lo lejos que estaba aún de los resultados. Se marchó a reunirse con los otros, mientras yo me quedé mirando las placas que quedaban. Para mi satisfacción, encontré un candidato supresor.
Poco más tarde tenía tres mutantes supresores que, por fortuna, distaban bastante entre sí a lo largo del mapa. Aislé los supresores y procedí a mapearlos. Mi teoría quedó refutada inmediatamente. En vez de encontrar cada supresor situado en el distante lugar previsto sobre el mapa, encontré que cada supresor estaba muy cerca del otro. El efecto de supresor tenía que deberse a otra razón.
Sin que yo lo supiera, otros investigadores se habían dado cuenta de que un mutante en rII podía tener un supresor en el mismo gen. Quizás el ejemplo más notable ocurrió en el Cal Tech. El físico teórico Dick Feynman se había interesado lo suficiente en estos problemas genéticos como para decidirse a hacer algunos experimentos. Encontró un ejemplo de un supresor interno. Ya que no sabía lo que esto podía implicar, se lo preguntó a su mentor Max Delbrück. Max sugirió que el mutante original había producido un aminoácido cambiado y que el segundo mutante lo había cambiado a otro aminoácido en otro lugar de la proteína, lo cual compensaba de alguna forma el primer cambio. Era fácil ver que esto podía suceder, pero no podía esperarse que fuera muy normal.
Por cierto, yo era muy consciente de esta posibilidad, pero no estaba satisfecho con ella, en parte porque ya tenía un conocimiento muy detallado de lo poco que se conocía entonces sobre la estructura de las proteínas. Decidí tratar de ver cuántos supresores diferentes podía tener un mutante determinado. Yo había seleccionado uno de los tres para estudiarlo más a fondo y evidentemente escogí aquel cuyo supresor se encontraba lo más lejos posible del mutante original, esperando que esto diera más juego. También me di cuenta de que dos de mis tres mutantes habían sido producidos por la proflavina. Aunque esto no tenía ningún significado estadístico, parecía interesante.
Para entonces ya tenía un poco más de experiencia, por lo que los experimentos fueron saliendo bastante deprisa. La genética de los fagos tiene la ventaja de que los experimentos son bastante rápidos una vez que se han puesto en marcha. No lleva mucho tiempo producir algunos centenares de cruces, ya que la manipulación es fácil y los cruces reales duran únicamente unos veinte minutos, que es el tiempo necesario para que un fago infecte la bacteria y se multiplique dentro de ella (intercambiando material genético en el proceso), haciendo que la célula se abra y muera. Los resultados de los cruces pueden ser entonces plaqueados sobre unas placas de Petri a las que se ha añadido una pequeña capa de bacterias. Entonces las placas tienen que ser incubadas hasta producir una cepa de bacterias. Allí donde un fago único ha aterrizado e infectado una célula, crecerá una colonia de fagos matando las bacterias locales y formando un pequeño agujero claro (llamado calva) sobre la cepa de las bacterias que crecen en la superficie de la placa. Este proceso dura unas horas, por lo que uno tiene un pequeño respiro mientras se está realizando. Entonces se sacan las placas de Petri de la estufa a 37 grados y se examinan para ver si tienen calvas o no y de qué tipo las tienen. Las calvas interesantes son seleccionadas, es decir, se recogen unos cuantos fagos con un pequeño pedazo de papel o un palillo, se hacen crecer un poco más y el proceso se repite por segunda vez para asegurarse de que el stock de fagos es puro. Si se trabaja a un ritmo razonable, es posible completar un conjunto de cruzamientos en un mismo día y prepararse para un nuevo conjunto de experimentos al día siguiente.
Cuando los experimentos se fueron haciendo más interesantes, me di cuenta de que con una planificación cuidadosa era posible hacer dos conjuntos sucesivos de cruzamientos en el mismo día. Esto suponía empezar temprano por la mañana, irse a casa para comer, hacer más experimentos por la tarde, irse a casa para la cena y hacer el conjunto final después de la cena. Por suerte Odile y yo vivíamos entonces a poca distancia del laboratorio, un hermoso paseo a través del centro histórico de Cambridge, lo que me volvía el trabajo más agradable. De hecho, Odile comentó que jamás me había visto tan contento como en aquel período en el que dedicaba todo mi tiempo a hacer experimentos, pero ello puede deberse a que durante varias semanas los experimentos funcionaron a la perfección.
Pronto me di cuenta de que mi mutante inicial no tenía uno, sino varios supresores distintos, los cuales mapeaban muy cerca del mutante original. Decidí que debía llamar a cada uno con un nombre distinto. A menudo trabajaba todo el fin de semana, tomándome el lunes libre para que la cocina de nuestro laboratorio (en la que se lavaba todo y también se preparaban las placas de Petri) pudiera ponerse al día. Ocurrió que un fin de semana necesitaba un nombre nuevo y no había nadie alrededor. Los mutantes se llaman en general con una letra seguida por un número. De esta forma, P31 significa el mutante treinta y uno de la serie P, probablemente producido por proflavina. Desgraciadamente no logré acordarme de qué letras habían sido utilizadas y por tanto decidí llamar a mi mutante FC0, ya que estaba bastante seguro de que nadie había utilizado mis iniciales para nombrar a mutantes. Los nuevos supresores fueron llamados entonces FC1, FC2, y así sucesivamente. El uso de mis propias iniciales hizo que alguna gente me considerara engreído, pero la explicación real es que la memoria me falla muy a menudo.
Todos los nuevos supresores parecían bastante buenos, sin ningún tipo de mutaciones filtrantes. Me pregunté entonces: ¿por qué no comprobar si ellos también tienen supresores? Y de hecho algunos los tenían. Fui incluso un paso más allá y encontré supresores de los supresores de los supresores.
Por lo tanto, ¿qué estaba ocurriendo? Afortunadamente teníamos ya a mano las ideas correctas. Supongamos que un mensaje genético era leído (para producir una proteína) en pasos de tres bases cada vez, empezando por un punto particular del mensaje. Para hacerlo más claro tomemos un mensaje muy sencillo, que simplemente repite el triplete TAG una y otra vez:

los puntos indican que hay mensaje a la vez antes y después de esta secuencia. Se añaden comas entre cada triplete para demostrar en qué «fase» la secuencia tiene que ser leída. Supuse que esta fase estaba determinada por una señal de «iniciación» especial, en algún lugar a la izquierda del fragmento que se muestra.
Supongamos que nuestro mutante original (ahora llamado FC0) ha añadido una base a la secuencia de bases. Entonces a partir de este punto la lectura estará fuera de paso (fuera de fase) y de esta forma producirá una proteína sin sentido, una proteína cuya secuencia de aminoácidos será completamente incorrecta y de esta forma el gen no podrá funcionar.
Nuestra secuencia simple ahora se habrá convertido en:

(La base que se ha añadido es una C para que el ejemplo sea claro, pero podría haber sido cualquiera de las otras bases).
Según esta interpretación, un supresor como el FC1 era la deleción de una base en un punto cercano. Entre FC0 y FC1 el mensaje sería incorrecto, ya que está leído en la fase que no corresponde, aunque en otros lugares la lectura sería correcta.
Nuestro ejemplo podría convertirse en:

o sea:

Si el trozo alterado en la secuencia de aminoácidos no era crucial (y en este caso había otra evidencia que así lo sugería), la proteína funcionaría bastante bien y el doble mutante (FC0 + FC1) podría comportarse de forma más parecida a uno de tipo silvestre que a un mutante sin filtraciones.
Por tanto, llamé – al primer conjunto de supresores. Al grupo siguiente, los supresores del primer conjunto de supresores, los llamé + y a sus supresores los llamé –.
Había comenzado los experimentos a principios de mayo y entonces el verano estaba avanzado. Previamente había decidido llevar a mi familia a veranear, y serían las primeras vacaciones propiamente dichas que tomaríamos, pues entonces mi posición económica había mejorado. Por una reducida suma, habíamos alquilado una gran casa sobre la vieja montaña de Tánger, una ciudad al norte de Africa, justo frente a Gibraltar. Ahí vivíamos con gran esplendor, teníamos un sirviente árabe viviendo en la casa y otro que venía todos los días. Odile y nuestra chica au pair alemana, Eleanora, aprendieron pronto a comprar regateando en el mercado árabe. Nuestras dos hijas mejoraban su natación en la playa mientras yo me pasaba el día en la terraza, a la sombra de las palmeras.
En nuestro camino hacia Tánger asistí a una reunión científica. Incluso en aquellos tiempos a los científicos no les gustaba ir a reuniones a menos que fuera en un lugar interesante. Esta reunión se daba en el Col de Voz, a medio camino del Mont Blanc. Expliqué mis resultados preliminares, los que fueron publicados poco después como una breve relación de la reunión.
Tras un mes en Tánger fui al Congreso de Bioquímica de Moscú de 1961, dejando a mi familia feliz en la gran casa durante una semana más. Moscú era muy diferente a la ciudad que viera en mi primera visita en 1945, durante la guerra. Ahora era verano en lugar de pleno invierno y todo parecía más brillante y próspero que en los oscuros días de la guerra. Me alojé en una habitación de estudiantes de la universidad, donde se celebraban las reuniones, y llegué a conocer a algunos huéspedes rusos. Una figura dominante era Igor Tamm, el físico ruso. La influencia de Lysenko, el hombre que durante un período había ahogado la genética en la Unión Soviética, estaba entonces menguando. Sentí que su eclipse era debido en gran medida al trabajo de físicos como Tamm, quienes tenían una influencia política considerable y sabían reconocer el sinsentido científico de lo que veían. Varios de nosotros fuimos invitados a dar charlas en la sección biológica del centro de Investigación de Energía Atómica ruso, algo que no habría podido ocurrir unos años antes. Dimos nuestras charlas en inglés, pero fueron brillantemente traducidas (por partes, según íbamos avanzando) por Bressler, un científico ruso que yo había conocido cuando visitó Cambridge. Bressler no sólo entendía lo que estaba diciendo sino que en algunos casos, como pude notar escuchándole, completaba las «referencias» que los conferenciantes iban dando, algo verdaderamente extraordinario.
La reunión de Moscú fue especialmente interesante gracias a los resultados presentados por Marshall Nirenberg, quien hasta entonces era bastante desconocido. Había rumores sobre estos experimentos, pero se desconocían los detalles. Matt Meselson, al que encontré en el corredor, me alertó sobre la charla que Marshall estaba dando en una remota sala de seminarios. Me impresionó de tal manera que pedí a Marshall que participara en una reunión mucho más amplia que yo presidía. Lo que él había descubierto era que se podía añadir un mensajero artificial a un sistema de un tubo de ensayo, que sintetizaba proteínas y conseguir que produjera alguna síntesis. Había añadido poli U (el RNA mensajero que consistía enteramente en la secuencia de uracilos) al sistema y éste sintetizaba fenilalanina. Esto sugería que UUU (suponiendo un código de tripletes) era el codón de fenilalanina (uno de los «veinte mágicos» aminoácidos), y realmente es así. Más tarde declaré que la audiencia había quedado «sobrecogida» (creo que originalmente escribí «electrizada») al recibir estas noticias. Seymour Benzer contradijo esta afirmación con una fotografía en la que todos parecían mortalmente aburridos. Sin embargo, fue un descubrimiento que marcó toda una época: tras él ya no se podía volver a mirar hacia atrás.
Durante esa semana en Moscú, incluso disfruté de alguna vida social. Me gustó visitar un apartamento al viejo estilo, con muebles pesados y una cama al lado de una larga librería, y también otro más moderno, con un tono mucho más ligero. El propietario de este último coleccionaba arte ruso moderno. Me divirtió ver cómo Alex Rich estaba enseñando un extraño baile norteamericano a nuestro anfitrión, un baile que más tarde supe que se llamaba twist. Como la cintura de Alex no es muy estilizada, el twist que enseñaba resultaba un tanto extraño.
Volví a Cambridge. El paso siguiente era realizar más experimentos para demostrar la validez de la idea de que tenía sentido denominar cada uno de nuestros mutantes rII como + o –. La teoría predecía que cualquier combinación del tipo (+ +) o (– –) sería un mutante. Mis colegas y yo construimos algunos de estos pares y tal como se había previsto eran mutantes sin filtraciones. La teoría predecía también que cualquier combinación del tipo (+ –) sería del tipo silvestre o próximo a él. Desde luego, sabíamos que esto sería verdad en algunos casos, ya que de esta forma habíamos encontrado el supresor en primer lugar, pero que muchas otras combinaciones (de + con –) debían ser probadas. Estas se llamaban «tíos y tías» ya que, para crearlas, a menudo debíamos poner juntos a un mutante de una generación con un mutante de una generación anterior pero de una procedencia diferente de la que él descendía. Había pedido a Sydney que se ocupara de que se intentaran algunas de éstas mientras yo estaba fuera, pero él tenía otras ideas y por tanto cuando volví tuve que hacerlo yo mismo.
En este punto apareció una pequeña complicación. Algunas de las combinaciones (+ –) que se preveían como de tipo salvaje resultaron ser un mutante. Esto lo explicamos suponiendo que en algunos casos el cambio local de fase entre la + y la – producía un mutante «sin sentido». Ahora sabemos que estos puntos sin sentido eran debidos a un triplete que terminaba la cadena polipeptídica y por tanto producía un fragmento de proteína no funcional. También me di cuenta de que esto dependía de la fase precisa de lectura. Para un código de tripletes no solamente hay una única fase correcta sino también dos fases incorrectas y por tanto la combinación (+ –), esto es, + seguido por – sería lógicamente diferente de una combinación (– +).
Volviendo a nuestro sencillo ejemplo, una combinación (+ –) podría ser:

Y una combinación (– +):

El primero tiene GTA entre las dos alteraciones; el segundo tiene AGT. Demostramos que nuestros fallos (+ –) o (– +) obedecían a esta regla, lo cual nos dio mucha confianza en que nuestras ideas estaban en la línea correcta.
Con anterioridad, Sydney había tenido una idea. Pensó que un mutante (+ –) podía volver a mutar hacia atrás dando un tipo silvestre. Probó uno, pero la mutación hacia atrás debía de haberse producido demasiado cerca de la existente y no pudo separarla. Una aproximación algo más laboriosa fue construir un mutante triple de la forma (+ + +) o (– – –). De acuerdo con estas ideas, éstos deberían ser del tipo silvestre, ya que los tres cambios sucesivos de la fase deberían restaurar la fase correcta suponiendo siempre, desde luego, que era un código de tripletes.
Para nuestra sencilla secuencia, un ejemplo podría ser:

Una vía directa pero laboriosa de construir un mutante triple consiste en escoger tres mutantes no muy alejados entre sí y todos +, y después otros dos pares, cada uno de los cuales tiene un mutante común en medio.
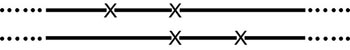
Figura 12.2. Cada línea representa una de las dos hebras progenitoras. Cada X representa una mutación. Es imposible recombinar las dos hebras progenitoras para dar una hebra que no tenga ninguna mutación. Siempre estará presente la mutación central. Además, parte de la progenie podrá tener las tres deleciones sobre la misma hebra.
Esta es la parte más ardua del trabajo, ya que no hay manera de seleccionar una combinación de mutantes como la deseada. Se tiene que hacer el cruce, y probar laboriosamente si el resultante tiene el fenotipo mutante, separando cada uno de ellos hasta que se encuentra el que es realmente el (+ +) que se está buscando. El paso final es fácil. Se cruzarán simplemente los dos dobles entre sí. Ya que cada uno contiene el mutante intermedio de los tres, no hay manera de que el cruce produzca un silvestre verdadero. Si del cruce aparecen placas aparentemente silvestres, es muy probable que sean las + + + que se habían buscado. En cualquier caso, es bastante fácil comprobar que este es el caso separando el supuesto triplete.
Desde luego el triplete sólo aparecería como un silvestre si el código es de tripletes. Si las bases fueran leídas en grupos de cuatro o cinco cada vez, lo cual, por lo que nosotros sabíamos, no era imposible, el (+ + +) sería un mutante y deberíamos construir un (+ + + +) o incluso un (+ + + + +). No todos en el laboratorio pensaban que el experimento funcionaría. Yo estaba casi seguro de que sí. También lo estaba Sydney, que por entonces se encontraba en París. Había hecho una lista de las tres posibles combinaciones (+ + +) para probar, pero después de que él se marchara, me di cuenta de que dos de ellas probablemente no funcionarían, debido a que producirían un terminador de cadena, por lo tanto construimos una tercera que parecía estar libre de esta complicación.
Por aquel entonces yo había pedido a Leslie Barnett que me ayudara. Los cruces finales se cumplieron debidamente y pusimos la pila de placas de Petri en la estufa. Volvimos después de la cena para inspeccionarlas. Una mirada a la placa decisiva fue suficiente. ¡Había en ella algunas calvas! El triple mutante tenía un comportamiento (un fenotipo) silvestre. Con cuidado comprobamos los números de las placas de Petri, para estar seguros de que habíamos mirado en la correcta. Todo estaba en orden. Miré a Leslie. «¿Te das cuenta», le dije, «de que tú y yo somos las únicas personas en el mundo que saben que hay un código de tripletes?»
El resultado, en cualquier caso, era importante. Nosotros teníamos tres mutantes distintos, cada uno de los cuales destruye la función del gen. Con ellos habíamos construido tres posibles mutantes dobles. Cada uno de éstos también hacía que el gen no funcionara. Pero si disponíamos los tres juntos en el mismo gen (hicimos experimentos separados para demostrar que estaban en el mismo virus y no en otro o en el resto de otros virus separados), el gen se ponía a funcionar otra vez. Esto resultaba fácil de entender si los mutantes eran adiciones o deleciones y si el código era en realidad un código de tripletes. En pocas palabras, habíamos encontrado la primera evidencia convincente de que el código era un código de tripletes.
He exagerado un poco. La evidencia también podría ser coherente en un código con seis bases en cada codón, pero esta posibilidad, como demostraron otros experimentos posteriores, era muy improbable y apenas podía tomarse en serio.
Todavía quedaba mucho trabajo para terminar con estos resultados. No construimos uno, sino seis diferentes triples (cinco del tipo + + + y uno del tipo – – –) y demostramos que todos se comportaban como los del tipo salvaje. En aquel momento estaba aún más ocupado que antes, a pesar de que Leslie era de gran ayuda. Y no es que no hubiera distracciones. Una noche, después de cenar, estaba trabajando en el laboratorio cuando una encantadora amiga mía apareció y se puso detrás de mí mientras yo continuaba manipulando los tubos y las placas. «Acompáñame a una fiesta», me dijo, pasando sus dedos por mi cabello. «Estoy demasiado ocupado», le contesté, «pero ¿dónde es?». «Bien», dijo, «creo que en tu propia casa». Finalmente llegamos a un acuerdo. Ella y Odile organizarían una pequeña fiesta y yo me reuniría con ellas cuando terminara de trabajar.
En retrospectiva, me parece impresionante lo poco que habíamos trabajado (había estado fuera durante seis semanas en el verano de mis viajes a Mont Blanc, Tánger y Moscú) y a la vez lo intensamente y deprisa que lo habíamos hecho. Yo había empezado el experimento clave en mayo. Sin embargo, el artículo fue publicado en Nature en el último número del año.
No nos detuvimos ahí. Sydney realizó por su cuenta muchos experimentos con el sistema. Más tarde decidimos que más valdría que publicásemos un informe completo de todo ello, por lo que Leslie Barnett y yo trabajamos muy duramente para atar todos los cabos sueltos que quedaban. Esto dio lugar a una consecuencia importante. Por entonces se sabía que los tripletes UAA y UAG eran terminadores de la cadena. Yo estaba convencido de que el UGA era el tercero. Sydney había pensado una vía muy complicada para comprobarlo genéticamente y los experimentos siempre decían que no era así. Cuando nos pusimos a escribir los resultados, nos dimos cuenta de que no habíamos realizado todos los experimentos posibles en este sentido. Para no dejar una brecha en una de nuestras tablas, pedimos a Leslie que como rutina hiciera los únicos que nosotros no habíamos llevado a cabo. Para nuestra sorpresa, el experimento funcionó. Repetimos los anteriores y esta vez también funcionaron. Resultó que lo que se había llevado a cabo en primer lugar había incluido un conjunto de controles para asegurarnos de que todo era como debía ser. Lamentablemente, en cada experimento un control u otro no había funcionado. Cuando todos funcionaron correctamente, el experimento sugirió claramente que la UAG era un terminador de la cadena.
Teníamos pensado dar a nuestros resultados un entierro decente en las augustas páginas de Phylosophical Transactions of The Royal Society. Como ahora habíamos dado con un resultado de cierto interés, sacamos nuestros experimentos del propuesto artículo del Phylosophical Transactions y los introdujimos en un artículo separado que apareció poco después en Nature. Quedé algo sorprendido al encontrar mi nombre en el borrador del artículo, ya que las normas de nuestro laboratorio consistían en que nadie ponía su propio nombre en un artículo a menos que hubiera hecho una contribución significativa. Un simple consejo amistoso no era suficiente. «¿Por qué habéis añadido mi nombre?», pregunté a Sydney. Me sonrió: «Por tu persistente hostigamiento», dijo, y yo dejé las cosas así.
Uno de los experimentos más laboriosos que Leslie realizó fue poner seis + juntos en un gen y demostrar que el resultado daba un fenotipo silvestre. Es difícil explicar lo tedioso y complicado que es este experimento. Requería seis + + + + + + juntos, uno tras otro, probando cada paso para comprobar que el gen en realidad tiene la estructura que se supone que tiene. Cuando la combinación final se ha producido y comprobado, debe ser separada paso a paso para asegurarse de que es lo que pensamos que era. Incluso una descripción preliminar de todo lo que Leslie llevó a cabo ocupó varias de las grandes páginas de Phylosophical Transactions.
Cuando estábamos repasando el manuscrito final le dije a Sydney que sospechaba que él y yo éramos las únicas personas en el mundo que lo leeríamos con cuidado. Para divertirnos decidimos añadir una referencia falsa, por lo que en un punto dado pusimos «Leonardo da Vinci (comunicación personal)» y lo presentamos a la Royal Society. Un lector (desconocido) lo pasó sin ningún comentario, pero recibimos una llamada telefónica de Bill Hayes, el otro lector, quien preguntó: «¿Quién es este joven italiano que trabaja en vuestro laboratorio?». No tuvimos más remedio que eliminarlo.
La demostración, por métodos genéticos, de que el código era un código de tripletes fue un tour de force, pero poco más tarde fue establecido también por métodos bioquímicos directos. A largo plazo tuvo mayor importancia la demostración de que los mutantes de acridina producían pequeñas deleciones y omisiones. Incluso esto no era insospechado, desde que Leonard Lerman había producido la evidencia químico-física de que las acridinas se colocaban entre las bases del DNA y ello podía fácilmente llevar a adiciones o deleciones del DNA cuando se copiaba. Además, la teoría tenía que quedar fehacientemente confirmada por métodos bioquímicos directos. Los bioquímicos Bill Dreyer y George Streisinger proyectaban hacerlo, aunque fueron algo lentos en la respuesta (en aquel tiempo era técnicamente difícil trabajar en bioquímica). Todos los meses Sydney y yo debatíamos si deberíamos ponernos a hacerlo nosotros, pero nos costó mucho decidirnos, porque George era un «viejo compañero» lo que significa que había pasado un tiempo en nuestro laboratorio. Finalmente George encontró la respuesta trabajando, no en los productos desconocidos de los dos genes, sino en la lisozima de los fagos. Ocurrió exactamente lo que esperábamos. Entre los mutantes se había alterado una cadena de aminoácidos y, además, encajaba bien con lo que se conocía del código genético y que justamente estaba empezando a aparecer.
Poco después, estuve en una reunión en Villa Serbelloni, junto al Lago Como, organizada por el biólogo Conrad Waddington (Wad para los amigos). Allí conocí al matemático René Thom. Lo primero que me dijo fue que nuestro trabajo con mutantes de acridina tenía que ser erróneo. Como acababa de enterarme de que nuestras ideas habían sido confirmadas por métodos bioquímicos, quedé algo sorprendido y le pregunté por qué pensaba así. Me explicó que si uno hacía, digamos, un mutante triple, se tenía necesariamente una distribución de Poisson de mutantes únicos, dobles, cuádruples, etc., y por tanto muchos argumentos no eran acertados. Ya que nosotros habíamos juntado nuestros mutantes con mucho trabajo (y lo habíamos comprobado con mucho cuidado), comprendí inmediatamente que esa objeción no tenía ninguna fuerza, ya que se basaba en un malentendido. O él no había leído nuestro artículo atentamente o, si lo había leído, no lo había entendido. Pero sé por experiencia que muchos matemáticos son intelectualmente perezosos y que les desagrada en especial leer artículos experimentales.
René Thom me dio la impresión de ser un buen matemático, aunque algo arrogante y nada dispuesto a explicar sus ideas en términos que los no matemáticos pudieran entender. Por suerte también estaba en la reunión otro topólogo, Christopher Zeeman, quien contribuyó especialmente a rechazar las ideas de Thom.
También tuve la impresión de que Thom entendía realmente muy poco sobre la forma de trabajar en la ciencia. Lo que entendía no le gustaba y se refería a ello con desprecio como «anglosajón». Me pareció que tenía profundas intuiciones biológicas, pero por desgracia de signo negativo. Sospeché que cualquier idea biológica que él pudiera tener sería probablemente errónea.