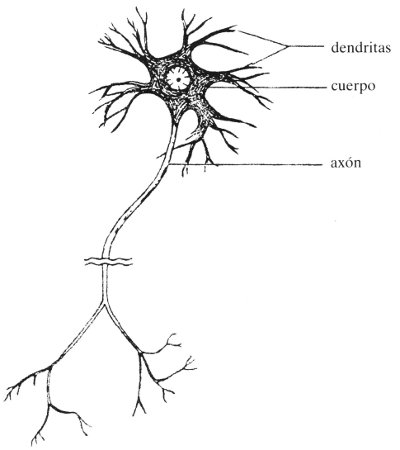
FIGURA 65. Dibujo esquemático de una neurona. [Adaptado de: D. Wooldridge, The Machinery of the Brain (New York: McGraw-Hill, 1963), p. 6.]
Cerebro y pensamiento
SÓLO CON EL ADVENIMIENTO de las computadoras comenzaron los intentos de producir máquinas de “pensar”, y tuvieron lugar caprichosas variantes acerca del tema del pensamiento. Se idearon programas cuyo “pensamiento” era, al pensamiento humano, lo que es a la locomoción humana uno de esos resortes que bajan escaleras saltando e invirtiéndose de un peldaño al siguiente. Súbitamente, las peculiaridades, las debilidades y las potencialidades, las extravagancias y las vicisitudes del pensamiento humano cayeron bajo la alusión de la recién descubierta capacidad de experimentar con formas artificiales, artesanales todavía, de pensamiento, o de aproximaciones al pensamiento. En los últimos veinte años, poco más o menos, hemos adquirido, como resultado, un género nuevo de perspectiva a propósito de lo que es y no es el pensamiento. Entretanto, los estudiosos del cerebro avanzaron mucho en el conocimiento del hardware de pequeña y gran escala de aquél. Si bien no obtuvieron un esclarecimiento considerable del modo en que el cerebro manipula los conceptos, sí han aportado algunas ideas sobre los mecanismos biológicos donde se asienta la manipulación de los pensamientos.
En este capítulo y en el siguiente, trataremos de vincular ciertas nociones surgidas de experimentaciones en el campo de la inteligencia de computadora, con algunos hechos brindados por ingeniosos ensayos con cerebros de animales vivos, y también con los resultados de las investigaciones dedicadas por los psicólogos del conocimiento a los procesos del pensamiento humano. El escenario ya ha sido levantado por el Preludio y Furmiga; ahora, desarrollaremos esas ideas más profundamente.
El pensamiento debe depender de la representación de la realidad en el hardware del cerebro. En los capítulos anteriores, hemos expuesto sistemas formales cuyo simbolismo representa dominios de la realidad matemática. ¿En qué grado es razonable utilizar tales sistemas formales como modelos de la forma en que el cerebro manipula ideas?
En el sistema mg, y luego en otros sistemas más complicados, vimos cómo la significación, en un sentido limitado del término, surge como resultado de un isomorfismo que hace corresponder a los símbolos tipográficos con números, operaciones y relaciones, y a las cadenas de símbolos tipográficos, con enunciados. Ahora bien, en el cerebro no hay símbolos tipográficos; sin embargo, hay algo mejor todavía: elementos activos que pueden almacenar información y transmitirla, además de recibirla de otros elementos activos. Por consiguiente, tenemos allí símbolos activos en lugar de símbolos tipográficos pasivos. En el cerebro, las reglas están entremezcladas con los propios símbolos, en tanto que, en el papel, los símbolos son entidades estáticas y las reglas están en nuestra cabeza.
Es importante evitar la suposición, sugerida por la naturaleza bastante estricta de todos los sistemas formales que hemos visto, de que el isomorfismo existente entre símbolos y cosas reales es una correspondencia rígida, elemento por elemento, como los hilos que unen un títere a la mano que lo guía. En TNT, la noción “cincuenta” puede ser expresada en diferentes formas simbólicas; por ejemplo,
((SSSSSSSO·SSSSSSSO)+(SO·SO))
((SSSSSSSSSSSO)+(SSSSSO·SSSSSO))
El hecho de que ambas expresiones representan el mismo número no es algo que esté en claro a priori. Podemos estar manipulándolas independientemente y, en algún punto, toparnos con un teorema que nos haga exclamar, “Oh… ¡Es ese número!”.
Nuestra mente puede albergar diferentes descripciones mentales de una misma persona; por ejemplo:
La persona cuyo libro envié a un amigo en Polonia, tiempo atrás.
El desconocido que trabó conversación conmigo y con mis amigas esta noche, en este café.
El hecho de que ambas expresiones aludan a la misma persona no es algo que esté en claro a priori. En algún momento, durante la velada, podemos toparnos con un tema de plática que nos conduzca a la revelación de que aquéllas designan a la misma persona, lo cual nos hará exclamar, “Oh… ¡Usted es esa persona!”.
No todas las descripciones de una persona tienen que ser relacionadas con determinado símbolo central de la misma, que almacene el nombre correspondiente. Las descripciones pueden ser elaboradas y manipuladas por sí mismas. Podemos inventar seres mediante el recurso de imaginar sus descripciones; podemos combinar dos descripciones si encontramos que ambas representan una entidad única; podemos seccionar una descripción en dos si encontramos que la misma representa dos cosas en lugar de una; y así sucesivamente. Este “cálculo de descripciones” está en el corazón del pensamiento; se ha dicho que es intensional, y no extensional, lo que significa que las descripciones pueden “flotar”, sin ser adheridas a objetos específicos, conocidos. La intensionalidad del pensamiento está conectada a su flexibilidad: nos da la capacidad de imaginar mundos hipotéticos, amalgamar entre sí descripciones distintas o partir una descripción en piezas separadas, etc.
Supongamos que una amiga, a quien le hemos prestado nuestro automóvil, nos telefonea para decimos que el coche patinó en un camino montañoso, golpeó contra algún borde y se dio la vuelta, y que ella ha escapado de la muerte de milagro. Una serie de imágenes se van suscitando en nuestra mente, las cuales se hacen progresivamente más vívidas en la medida en que nuestra amiga agrega mayores detalles, hasta que el final “lo vemos todo con los ojos de nuestra mente”. Entonces, ella nos dice que todo ha sido una broma del Día del Inocente y que ella y el coche están perfectamente bien. En muchos sentidos, esta última circunstancia no es pertinente: la anécdota y las imágenes no pierden nada de su vividez y su recuerdo se retiene durante mucho, muchísimo tiempo. Más tarde, es posible inclusive que pensemos de nuestra amiga que es una conductora insegura, todo a causa del vigor de la primera impresión, la cual debería haber sido barrida cuando supimos que ella no decía la verdad. La fantasía y los hechos se entremezclan muy íntimamente en nuestras mentes, y ello es debido a que el pensamiento abarca la elaboración y la manipulación de descripciones completas, las cuales no necesitan, en absoluto, ser ligadas a sucesos o a cosas reales.
Una representación flexible, intensional del mundo: en eso consiste el pensamiento, por donde se lo mire. Ahora bien, ¿cómo es posible que un sistema fisiológico como el cerebro dé soporte a un sistema como el del pensamiento?
Las células más importantes del cerebro son las células nerviosas, o neuronas (véase la figura 65), cuyo número es de aproximadamente diez mil millones. (Curiosamente, exceden con mucho a éstas en número —su proporción es de casi diez por una— las neuroglias. Se cree que las glías llenan más de una función de colaboración con el papel estelar que desempeñan las neuronas, por lo que no hablaremos de ellas ahora). Cada neurona tiene una cantidad de sinapsis (“puertas de entrada”) y un axón (“canal de salida”). La entrada y la salida son flujos electroquímicos: es decir, iones en movimiento. Entre las puertas de entrada y el canal de salida de una neurona se encuentra el cuerpo de la célula: donde se toman las “decisiones”. El tipo de decisión que enfrenta —lo cual tiene lugar hasta mil veces por segundo— es el siguiente: si se excitan o no; excitarse significa liberar iones por su axón, los cuales finalmente cruzarán las puertas de entrada de una o más neuronas, haciendo que éstas se enfrenten a una alternativa similar. La decisión se adopta de una manera muy simple: si la suma de todas las entradas sobrepasa un determinado umbral, sí; en caso contrario, no. Algunas de las entradas pueden ser entradas negativas, las cuales suprimen las entradas positivas de otras procedencias. En cualquier caso, es la mera suma lo que rige los niveles más bajos de la mente. Podemos parafrasear la célebre expresión de Descartes: “Pienso, luego sumo” (del latín Cogito, ergo soy).
FIGURA 65. Dibujo esquemático de una neurona. [Adaptado de: D. Wooldridge, The Machinery of the Brain (New York: McGraw-Hill, 1963), p. 6.]
Ahora bien, a pesar de que la manera de adoptar la decisión parece muy sencilla, hay un hecho que la complica: puede haber nada menos que 200.000 puertas de entrada distintas en una neurona, lo cual significa que pueden estar comprendidos hasta 200.000 sumandos en la determinación de la próxima acción de la neurona. Ya tomada la decisión, un impulso de iones se dirige hacia el extremo terminal del axón. Antes de llegar allí, los iones pueden encontrarse con una o varias bifurcaciones. En este caso, el impulso unitario de salida se divide para recorrer también la bifurcación y, en el momento de alcanzar el extremo, “la” se ha convertido en “las”, y es posible que no arriben al mismo tiempo porque las ramas del axón por las cuales viajan pueden tener diferentes longitudes y presentar diferentes grados de resistencia. Lo más importante, no obstante, es que todas comienzan como un solo impulso que se desplaza desde el cuerpo de la célula y en dirección al exterior. Después de excitarse, una neurona necesita un corto lapso de recuperación para excitarse de nuevo; lo habitual es que ello le insuma un milisegundo, de modo que una neurona puede excitarse hasta mil veces por segundo.
Acabamos de describir las “hormigas” del cerebro. ¿Y qué diremos de los “equipos” o “señales”? ¿Y de los símbolos? Hacemos la siguiente observación: a pesar de la complejidad de su entrada, una neurona individual únicamente puede responder en una forma primitiva, es decir, excitándose o no excitándose. Esto implica una cantidad muy reducida de información. Indudablemente, para el traslado o el procesamiento de grandes volúmenes de información deben intervenir muchas neuronas. En consecuencia, uno puede conjeturar que deben existir estructuras mayores, compuestas por muchas neuronas, encargadas de manejar conceptos en un nivel más alto. Por cierto que verdaderamente es así, pero la concepción ingenua según la cual hay un grupo fijo de neuronas por cada concepto es, casi con seguridad, falsa.
Hay muchas porciones anatómicas del cerebro que pueden ser distinguidas entre sí, tales como el encéfalo, el cerebelo, el hipotálamo (véase figura 66). El encéfalo es la más grande de estas partes, y está dividido en dos hemisferios: derecho e izquierdo. Los escasos milímetros exteriores de cada uno de los hemisferios cerebrales están revestidos por una “cáscara” estratificada, o corteza cerebral. La cantidad de corteza cerebral es el principal rasgo de diferenciación, desde el punto de vista anatómico, entre los cerebros humanos y los de especies menos inteligentes. No describiremos en detalle ninguno de los subórganos cerebrales porque, en función de lo que sabemos hasta la fecha, sólo puede ser trazada la más rudimentaria de las correspondencias entre los subórganos de tan gran escala y las actividades, mentales o físicas, de las cuales son responsables. Por ejemplo, es sabido que el lenguaje tiene su centro en uno de ambos hemisferios, principalmente el izquierdo, por lo común. También, que el cerebelo es el lugar desde donde son enviadas series de impulsos a los músculos, con vistas al control de la actividad motora. Pero de qué modo estas áreas desempeñan sus funciones sigue siendo un gran misterio.
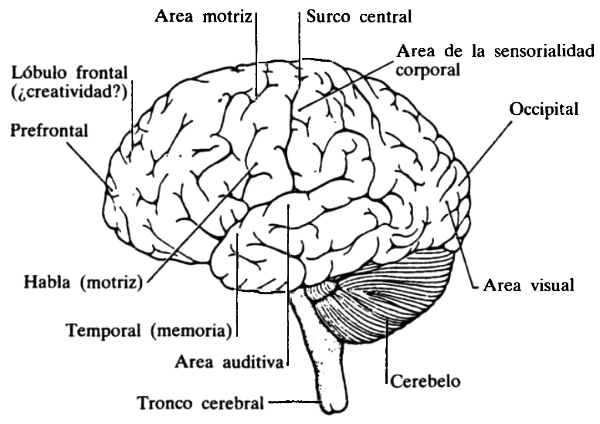
FIGURA 66. El cerebro humano, visto desde el perfil izquierdo. Es extraño que el área visual esté ubicada en la parte de atrás de la cabeza. [De: Steven Rose, The Conscious Brain, updated ed. (New York: Vintage, 1966), p. 50.]
Ahora bien, aquí aparece una pregunta en extremo importante. ¿Si el pensamiento tiene lugar en el cerebro, cuán diferentes, entonces, son dos cerebros entre sí? ¿Cuán diferente es mi cerebro del cerebro del lector? Sin duda, el lector no piensa del modo en que yo lo hago, ni del modo en que lo hace nadie. Pero todos tenemos las mismas divisiones anatómicas en nuestro cerebro. ¿Hasta dónde se extiende esta identidad entre cerebros? ¿Llega al nivel neuronal? Sí, en el caso de animales ubicados en un nivel bastante bajo de la jerarquía mental: la humilde lombriz de tierra, por ejemplo. La cita que sigue corresponde a un neurofisiólogo, David Hubel, y forma parte de palabras que pronunciara en una conferencia sobre comunicación con inteligencias extraterrestres:
El número de células nerviosas de un animal como la lombriz debe ser, creo, del orden de los miles. Algo muy interesante es que podemos señalar una célula individual en particular, en una determinada lombriz de tierra, y luego identificar la misma célula, la célula correspondiente, en otra lombriz de tierra de la misma especie.[1]
¡Las lombrices de tierra tienen cerebros isomórficos! Uno podría decir, “Hay solamente una lombriz de tierra”.
Pero tal correspondibilidad elemento por elemento entre cerebros individuales desaparece muy pronto cuando se comienza a ascender en la jerarquía mental y aumenta el número de neuronas… ¡lo cual confirma las sospechas de uno en el sentido de que no hay solamente un ser humano! Sin embargo, pueden ser detectadas considerables similitudes físicas entre diferentes cerebros humanos si son comparados dentro de una escala mayor que la de una neurona, pero menor que la de los subórganos principales del cerebro. ¿Qué denota esto con relación al modo en que las diferencias mentales individuales están representadas en el cerebro físico? Si examináramos las interconexiones de mis neuronas, ¿encontraríamos diversas estructuras de las que se pudiera determinar que codifican cosas específicas que conozco, creencias específicas que sostengo, esperanzas específicas, temores, preferencias y rechazos específicos que albergo? Si las experiencias mentales pueden ser atribuidas al cerebro, ¿se podrá, del mismo modo, establecer la localización específica del conocimiento y de otras esferas de la vida mental en el cerebro, o en subsistemas físicos específicos del cerebro? Ésta es una interrogación central, a la que retornaremos repetidas veces en el curso de este capítulo y del siguiente.
Preocupado por responder a esa misma interrogación, el neurólogo Karl Lashley intentó descubrir, por medio de experimentos que comenzaron alrededor de 1920 y se extendieron durante muchos años, en qué parte del cerebro almacena una rata los conocimientos que adquiere en materia de recorridos de laberintos. Steven Rose, en su libro The Conscious Brain, describe de la siguiente forma los ensayos y las tribulaciones de Lashley:
Lashley intentaba identificar el locus de la memoria dentro de la corteza; para conseguirlo, primero adiestró ratas para que atravesasen laberintos, y luego les extrajo diversas regiones corticales. A continuación, dejó que los animales se recuperaran, para después verificar si había retención de la habilidad adquirida. Lashley se sorprendió al descubrir que no era posible hallar una región en particular que correspondiese a la capacidad de recordar el camino seguido dentro de un laberinto. En cambio, descubrió que todas las ratas que habían sufrido la extracción de regiones corticales padecían de algún tipo de perturbación y que el alcance de ésta era aproximadamente proporcional a la cantidad de corteza extraída. La extracción de corteza había afectado las capacidades motrices y sensoriales de los animales: cojeaban, se valían de saltitos para avanzar, se bamboleaban, vacilaban, pero de alguna manera se las arreglaban, en todos los casos, para atravesar el laberinto. En cuanto a la memoria, aparentemente la relación de la corteza con ella era equipotencial, es decir, que todas las regiones presentaban la misma utilidad potencial. Por cierto, en su último trabajo, In Search of the Engram, aparecido en 1950, Lashley afirmó, más bien lúgubremente, que la única conclusión consistía en que localizar la memoria no era en absoluto posible.[2]
Es curioso que, aproximadamente al mismo tiempo que Lashley realizaba su último trabajo, hacia fines de la década de los cuarenta, en Canadá se desarrollaba una investigación que producía evidencias totalmente opuestas. El neurocirujano Wilder Penfield se dedicó a examinar las reacciones de pacientes cuyos cerebros habían sido operados. Se valió de la inserción de electrodos en diversas partes de dichos cerebros, los cuales estaban expuestos, y del envío de pequeños impulsos eléctricos para estimular la neurona o las neuronas a las cuales habían sido conectados los electrodos. Estos impulsos eran similares a los que provienen de otras neuronas. Penfield descubrió que la estimulación de ciertas neuronas generaba, sin excepción, imágenes o sensaciones específicas en el paciente. Estas impresiones artificialmente provocadas comprendieron desde temores extraños e indefinibles hasta sonidos y colores; y lo más llamativo de todo fue que incluyeron recuerdos de sucesiones enteras de hechos acontecidos en las más tempranas etapas de la existencia, como por ejemplo una fiesta infantil de cumpleaños. El conjunto de lugares que podía desencadenar tales hechos específicos era extremadamente pequeño: básicamente, se centraba en una sola neurona. Ahora bien, estos hallazgos de Penfield se oponen radicalmente a las conclusiones de Lashley, puesto que parecen trasuntar que las áreas locales tenían la responsabilidad de funciones a las cuales se puede considerar, al fin y al cabo, memorias específicas.
¿Cómo entender esto? Una explicación podría ser la de que esas memorias están codificadas localmente, pero repitiéndose incontables veces en diferentes áreas de la corteza: una estrategia así desarrollada a través de la evolución, quizás en previsión de posibles pérdidas de corteza en los combates, o en experimentos dirigidos por neurofisiólogos. Otra explicación sería la de que dichas memorias pueden ser reconstruidas a partir de procesos dinámicos que abarcan el cerebro en su conjunto, pero pueden ser desencadenadas a partir de puntos localizados. Esta teoría se basa en una noción sugerida por las redes telefónicas modernas, donde el itinerario de una comunicación de larga distancia no es predictible por adelantado, pues se lo establece en el momento en que se produce el requerimiento correspondiente y depende de la situación vigente en el sistema nacional entero. La destrucción de cualquier sector local de la red no impediría las comunicaciones; sólo provocaría su desviación en torno a las áreas dañadas. En este sentido, todo llamado es, potencialmente, no localizable. Sin embargo, toda llamada, precisamente, conecta entre sí dos puntos específicos; en este sentido, toda llamada es localizable.
Algunos de los más interesantes y significativos trabajos en materia de localización de procesos cerebrales son los realizados en Harvard, en el trascurso de los últimos quince años, por David Hubel y Torsten Wiesel. Consiguieron hacer corresponder los recorridos visuales de los cerebros de distintos gatos, comenzando por las neuronas de la retina, siguiendo sus conexiones hacia la parte de atrás de la cabeza, pasando a través del “puesto de relevo” de la articulación lateral, hasta terminar en la corteza visual, situada en la parte posterior del cerebro. Primero de todo, es notable que existan recorridos neurales bien definidos, teniendo en cuenta las conclusiones de Lashley. Pero más notables todavía son las propiedades de las neuronas localizadas en los diferentes hitos del recorrido.
Tenemos que las neuronas de la retina son, en primer lugar, sensores de contraste; más específicamente expresado, tal es la forma en que actúan. Cada una de esas neuronas se excita, normalmente, a “velocidad de crucero”. Cuando su porción de retina es herida por la luz, puede excitarse con mayor rapidez o con mayor lentitud, e inclusive cesar de hacerlo. Pero esto sucederá así solamente en el caso de que la parte circundante de la retina esté menos iluminada. Luego, esto significa que hay dos tipos de neuronas: “centros de encendido” y “centros de apagado”. Las neuronas centro de encendido son aquellas cuya frecuencia de excitación se incrementa en todos los sitios donde, dentro de la pequeña área circular de la retina con respecto a la cual son sensitivas, el centro está iluminado pero las inmediaciones a oscuras; las neuronas centro de apagado son aquellas que se excitan más rápidamente cuando el centro está en la oscuridad pero los contornos están iluminados. Si un patrón de los que rigen los centros de encendido es presentado a una neurona centro de apagado, ésta reducirá su frecuencia de excitación (y viceversa). Ambos tipos de neurona no experimentarán alteración alguna frente a una iluminación uniforme: continuarán excitándose a velocidad de crucero.
Desde la retina, las señales procedentes de estas neuronas se dirigen, a través del nervio óptico, a la articulación lateral, ubicada en algún punto próximo a la mitad del cerebro. Allí, es posible encontrar una proyección directa de la superficie retinal, en el sentido de que hay neuronas de la articulación lateral que son puestas en acción únicamente por estímulos específicos producidos en áreas específicas de la retina. En tal sentido, la articulación lateral es decepcionante: parece ser tan sólo un “puesto de relevo”, y no un nuevo procesador (la sensitividad de contraste hace su aporte a la articulación lateral, donde, aparentemente, es realizada). La imagen retinal es codificada de una manera directa por los patrones de excitación de las neuronas de la articulación lateral, a pesar del hecho de que las neuronas no están dispuestas allí sobre una superficie bidimensional, como lo es la forma de la retina, sino en un bloque tridimensional. Así, dos dimensiones son proyectadas sobre tres, sin que se altere la información contenida: un isomorfismo. Probablemente haya alguna profunda significación en el cambio que experimenta la dimensionalidad de la representación, el cual no es apreciado aún en forma completa. De cualquier modo, hay muchos otros estadios inexplicados de la visión que, en lugar de provocarnos frustración, deberían hacemos sentir complacidos puesto que, en alguna medida, ¡hemos resuelto lo referente a éste!
Desde la articulación lateral, las señales se dirigen a la corteza visual. Aquí ocurren ciertos tipos nuevos de procesos. Las células de la corteza visual se dividen en tres categorías: simples, complejas e hipercomplejas. Las células simples actúan de manera muy parecida a las retinales o a las de la articulación lateral: responden a puntos iluminados u oscuros, contrastados por sus respectivos contornos, en regiones particulares de la retina. Las células complejas, en cambio, son generalmente alimentadas por un centenar o más de otras células, y detectan las fajas de luz o de oscuridad orientadas en ángulos específicos de la retina (véase figura 67). Las células hipercomplejas responden a rincones, fajas e inclusive “lenguas” que se mueven en direcciones específicas (véase también la figura 67). Estas células son tan altamente especializadas que a veces se las denomina “células hipercomplejas de jerarquía más elevada”.
A causa del descubrimiento de células de la corteza visual que pueden ser accionadas por estímulos de complejidad cada vez mayor, hay quienes se han preguntado si las cosas no marchan en la dirección de “una célula, un concepto”: por ejemplo, tendríamos una “célula abuela” que se excitaría si, y solamente si, aparece nuestra abuela en nuestro campo de visión. Esta ilustración un tanto humorística de una presunta “célula superhipercompleja” no es considerada muy seriamente. Sin embargo, la teoría alternativa a ésta no impresiona como obviamente razonable. Una posibilidad consiste en que las redes neurales más extensas sean excitadas colectivamente por estímulos visuales suficientemente complejos. Por supuesto, el accionamiento de estas unidades multineuronales más extensas tendría que provenir, de alguna manera, de la integración de señales emanadas de muchas células hipercomplejas. Cómo puede ser que se produzca esto no lo sabe nadie. Precisamente cuando parecemos acercarnos al umbral donde las “señales” se convierten en “símbolos”, perdemos la pista: una angustiosa e interminable historia. Volveremos pronto a esta historia y trataremos de completarla en alguna de sus partes.
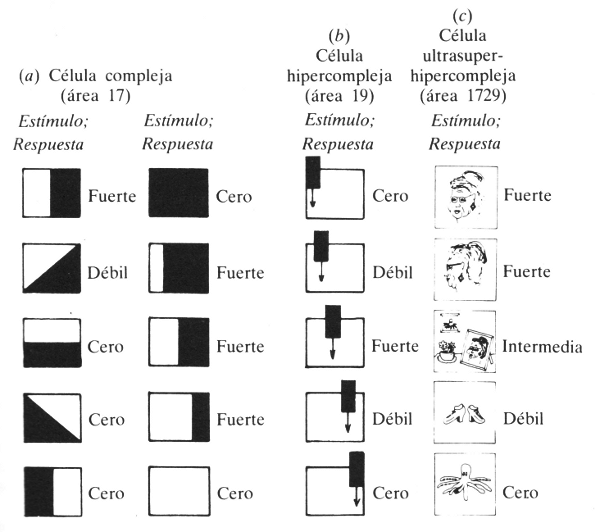
Figura 67. Respuestas de determinadas neuronas simples a sus patrones de estimulación.
(a) Esta neurona detectora de bordes busca bordes verticales que tengan luz a la izquierda y oscuridad a la derecha. La primera columna muestra que la orientación de un borde es pertinente para esta neurona. La segunda columna muestra que la posición del borde carece de pertinencia en el caso particular de esta neurona.
(b) Muestra cómo una célula hipercompleja responde más selectivamente: en este diagrama, sólo lo hace cuando la lengua descendente está en el centro del campo.
(c) Vemos las respuestas de una hipotética “célula abuela” a diversos estímulos arbitrarios; el lector puede entretenerse considerando cómo respondería a los mismos estímulos una “célula pulpo”.
Anteriormente, mencioné el isomorfismo de trama gruesa que existe, en el plano de una gran escala anatómica, entre todos los cerebros humanos, y el isomorfismo de trama muy fina, en el nivel neural, que existe entre los cerebros de las lombrices de tierra. Es sumamente interesante también la existencia de un isomorfismo entre los aparatos visuales del gato, del mono y del ser humano, isomorfismo cuya “trama” es de algún tipo entre grueso y fino. Veamos cómo funciona: en primer término, las tres especies han “destinado” áreas de corteza ubicadas en la parte posterior de sus cerebros para que allí tenga lugar el procesamiento visual: la corteza visual; en segundo término, la corteza visual de cada uno de ellos está dividida en tres subregiones, llamadas las áreas 17, 18 y 19 de la corteza. Estas áreas son universales, en el sentido de que pueden ser localizadas en el cerebro de cualquier individuo normal perteneciente a cualquiera de las tres especies. Dentro de cada una de las áreas, es posible ir aún más lejos, hasta llegar a la organización “columnaria” de la corteza visual. En forma perpendicular a la superficie de la corteza, y desplazándose radialmente hacia adentro, en dirección al cerebro interior, hallamos las neuronas visuales dispuestas en “columnas”: esto es, todas las conexiones se desplazan a lo largo de la dirección radial, columnaria; no lo hacen de una columna a otra. Y cada columna se corresponde con una pequeña y específica región de la retina. La cantidad de columnas no es la misma en cada individuo, por lo que no es posible encontrar “la misma columna”. Dentro de una de éstas, por último, hay estratos donde es más factible descubrir neuronas simples, y otros estratos donde es más factible descubrir neuronas complejas. (Las neuronas hipercomplejas tienden a ser descubiertas, predominantemente, en las áreas 18 y 19, en tanto que las simples y las complejas lo son, más que nada, en el área 17). Aparentemente, aquí se agotan los isomorfismos correspondientes a este nivel de detalle. Desde este punto hacia abajo, hasta el nivel neuronal individual, cada gato, mono o ser humano singular constituyen un ejemplar absolutamente único: algo tan peculiar como las huellas digitales o la firma autógrafa.
Una diferencia pequeña, pero quizás importante, que distingue el procesamiento visual de los gatos del de los monos está relacionada con el punto en el cual la información proveniente de ambos ojos se integra, para producir una sola señal combinada de más alto nivel. Esto tiene lugar ligeramente después en el mono que en el gato, lo cual da a la señal de cada uno de los ojos un lapso ligeramente más extenso para ser procesada por separado. No se trata de algo demasiado asombroso, pues es razonable que, cuanto más alto se ubique una especie en la jerarquía de la inteligencia, más complejos serán los problemas que su sistema visual sea llamado a responder; luego, las señales, en este caso, tendrán que atravesar más y más procesamientos previos, antes de recibir un “rótulo” definitivo. Esto es rotundamente confirmado por la observación de las facultades visuales de un ternero recién nacido, el cual pareciera llegar a la vida con la misma capacidad de discriminación visual que tendrá siempre: se apartará de las personas o de los perros, pero no de sus congéneres. Es probable que su sistema visual tenga completo, antes del nacimiento, su “hardware”, y que abarque relativamente pocos procesos corticales. En cambio, el sistema visual humano, tan profundamente afincado en la corteza, requiere varios años para alcanzar su madurez.
Una cosa desconcertante, a propósito de los descubrimientos efectuados hasta ahora acerca de la organización del cerebro, es que han sido encontradas muy escasas correspondencias directas entre la gran escala del hardware y el alto nivel del software. La corteza visual, por ejemplo, es una pieza en gran escala de hardware, destinada por entero a una clara finalidad del género software: el procesamiento de la información visual (pese a que todos los procesos descubiertos hasta el momento corresponden, sin excepción, al bajo nivel). En la corteza cerebral, no se ha localizado nada que se parezca al reconocimiento de objetos. Ello significa que nadie sabe dónde o cómo la salida de las células complejas e hipercomplejas es transformada en reconocimiento consciente de formas, lugares, imágenes, rostros, etc. Los estudios han tratado de encontrar pruebas de un “embudamiento” de muchas respuestas neurales de bajo nivel dentro de respuestas, progresivamente más escasas, de nivel mayor, que culminen en algo semejante a nuestra conocida célula abuela, o en alguna clase de red multineuronal, como la que ya mencionamos. Es evidente que ello no será hallado en grandes divisiones anatómicas que se atribuyan al cerebro, sino, por el contrario, en los resultados de análisis microscópicos.
Una explicación alternativa a la de la célula abuela podría consistir en suponer un conjunto fijo de neuronas: unas pocas docenas, digamos, en el extremo fino del “embudo”, todas las cuales se excitan cuando Abuelita ingresa al campo de visión. Y para cada diferente objeto reconocible habría una red única, y un proceso de embudamiento por el que se canalizaría la red. Hay alternativas más complicadas dentro de líneas similares, que presumen varios modos diversos de excitación por parte de las redes, en lugar de un solo modo fijo. Tales redes serían los “símbolos” de nuestro cerebro.
Ahora bien, ¿es necesario el embudamiento? Quizá cuando se observa un objeto sea identificado implícitamente por su “firma” en la corteza visual, lo cual implica la reunión de las respuestas de las células simples, complejas e hipercomplejas. Quizá el cerebro no necesite de ningún mecanismo adicional, aparte del recién mencionado, frente a una forma determinada. Esta teoría, sin embargo, plantea el siguiente problema: supongamos que estamos mirando un cuadro; éste registra su firma en nuestra corteza visual, ¿pero cómo haremos luego para extraer, de esa firma, una descripción verbal de la imagen? Por ejemplo, las pinturas de Edouard Vuillard, un postimpresionista francés, requieren a menudo algunos segundos de examen hasta que, de súbito, surge a la vista una figura humana. Es de suponer que la firma marca su impresión en la corteza visual durante la primera fracción de segundo, pero la pintura es comprendida sólo algunos segundos después. Esto no es más que un ejemplo aislado de un fenómeno muy habitual: la sensación de que algo se “cristaliza” en nuestra mente, en el momento del reconocimiento, lo cual tiene lugar no cuando los rayos de luz chocan nuestra retina, sino un poco más tarde, luego de que cierta parte de nuestra inteligencia ha tenido oportunidad de influir en las señales retinales.
La metáfora de la cristalización permite la bella imagen, derivada de la mecánica estadística, de una miríada de actividades microscópicas e inconexas en el interior de un medio, donde se crean, lentamente, regiones de coherencia que se diseminan y se agradan; por último, la miríada de pequeños hechos habrá realizado una completa reorganización estructural de su medio de abajo arriba, convirtiéndolo, de una reunión caótica de elementos independientes, en una estructura amplia, coherente, totalmente articulada. Si las actividades neurales iniciales son consideradas como independientes, y el resultado final de sus muchas excitaciones independientes como la concreción de un gran “módulo” bien definido de neuronas, se debe entender entonces que la palabra “cristalización” es perfectamente adecuada.
Otro argumento en favor del embudamiento se basa en la circunstancia de que hay una miríada de distintas escenas que pueden provocamos la impresión de que hemos percibido ya el mismo objeto: nuestra abuela, por ejemplo, quien puede estar sonriendo o con el ceño fruncido; con un chal sobre sus hombros, o no; en un jardín en flor o en una oscura estación de ferrocarril; vista desde cerca o desde lejos, de frente o de perfil, y así sucesivamente. Todas estas escenas producen firmas sumamente diferentes en la corteza visual, pero todas nos mueven a decir, “Hola, Abuelita”. De modo, pues, que un proceso de embudamiento debe tener lugar en algún momento posterior a la recepción de la firma visual y anterior al proferimiento de las palabras. Uno puede sostener que tal embudamiento no es parte de la percepción de Abuelita sino, tan sólo, parte de la verbalización. Pero semejante fragmentación del proceso impresiona como algo muy antinatural, pues es posible la utilización interna, sin verbalizarla, de la información de que se trata de Abuelita. Sería muy difícil manejar toda la información por entero en la corteza visual, cuando una parte tan grande de la misma es descartada, ya que no nos preocupamos por el lado hacia el cual cae la sombra en la escena, o por el número de botones de la blusa, etcétera.
Otra de las dificultades que presenta una teoría no embudadora es cómo explica la existencia de interpretaciones diferentes para una firma única: el cuadro Convexo y cóncavo, de Escher (figura 23), por ejemplo. Así como nos parece obvio que no percibimos simplemente puntos en una pantalla de televisión, sino bloques, suena ridícula la postulación de que la percepción tiene lugar cuando una gigantesca “firma” de apariencia punteada es producida en la corteza visual. Tiene que haber algún embudamiento, cuyo resultado final sea desencadenar ciertos módulos específicos de neuronas, cada uno de los cuales se asocie con los conceptos —los bloques— de la imagen.
Somos así llevados a la conclusión de que, para cada concepto, hay un módulo perfectamente bien definido, el cual ha podido ser desencadenado; un módulo que consiste en un reducido grupo de neuronas; un “complejo neural” del tipo comentado más atrás. Uno de los problemas que involucra esta teoría —por lo menos, en el caso de que se la asuma ingenuamente— es su posible sugerencia en el sentido de que se estaría en condiciones de localizar tales módulos en algún punto ubicado dentro del cerebro. Tal cosa no ha sido lograda todavía, y algunas de las comprobaciones efectuadas, como por ejemplo las surgidas de los ensayos de Lashley, se pronuncian en contra de la localización. Sin embargo, una afirmación concluyente al respecto sería prematura. Puede que existan muchas réplicas, diseminadas por todas partes, de cada módulo, o que éstos se encuentren físicamente superpuestos; cualquiera de ambas situaciones tendería a dificultar toda forma de separación de las neuronas en “paquetes”. Quizá los complejos se parezcan mucho a delgados panqueques y que constituyan, así, estratos compactos, los cuales se entrecruzarían de vez en cuando; quizá sea como largas serpientes que se enroscan entre sí, irguiéndose aquí y allá, como de cabezas de cobras; quizá sean como telarañas; o, quizá, circuitos donde las señales giren y giren describiendo formas más extrañas que las evoluciones de un mosquito antes de picar. No hay forma de saberlo. Es posible, inclusive, que estos módulos sean fenómenos del orden del software, y no del hardware, pero esto es algo de lo cual hablaremos más adelante.
Son muchas las preguntas que surgen acerca de estos hipotéticos complejos neurales. Por ejemplo:
¿Se extienden a las regiones más bajas del cerebro, tales como el mesencéfalo, el hipotálamo, etc.?
¿Una misma neurona puede pertenecer a más de uno de dichos complejos?
¿A cuántos de dichos complejos puede pertenecer una misma neurona?
¿A través de cuántas neuronas pueden darse las superposiciones de dichos complejos?
¿Estos complejos son prácticamente idénticos en todo el mundo?
¿Los complejos correspondientes se localizan en lugares correspondientes, en los cerebros de distintas personas?
¿Se superponen del mismo modo en el cerebro de todo el mundo?
Filosóficamente, la pregunta más importante es la siguiente: ¿Qué nos indicaría la existencia de módulos, la de —por ejemplo— un módulo abuela? ¿Ello nos permitiría penetrar en el fenómeno de nuestra conciencia? ¿O seguiríamos en la misma oscuridad en la que estamos ahora, pese a que ya sabemos que el cerebro está constituido por neuronas y neuroglias?
Tal como se puede conjeturar a través de la lectura de la Furmiga, me inclino a creer que es muy largo el camino por recorrer para aproximarnos a una comprensión del fenómeno de la conciencia. El paso crucial que imprescindiblemente se debe transitar es el que va desde una descripción —neurona por neurona— de bajo nivel del estado de un cerebro, a una descripción —módulo por módulo— de alto nivel del estado del mismo cerebro. O, para retornar a la sugestiva terminología utilizada en la Furmiga, necesitamos desplazar la descripción del estado cerebral desde el nivel de la señal, al nivel del símbolo.
A todos estos hipotéticos complejos neurales, módulos neurales, paquetes neurales, redes neurales, unidades multineuronales —llamémoslos como queramos, según que se nos aparezcan bajo la forma de panqueques, de jardines, de cacahuetes, de reptiles, de copos de nieve o inclusive de olas en relieve— los vamos a designar, desde este momento, símbolos. Ya se hizo alusión en el Diálogo a la descripción de un estado cerebral en función de los símbolos. ¿Cómo será una descripción así? ¿Qué clase de conceptos será razonable pensar que pueden ser “simbolizados”? ¿Qué clase de relaciones mantendrán los símbolos entre sí? ¿Y qué profundizaciones en el conocimiento de la conciencia suministrará esta imagen en su conjunto?
Lo primero por subrayar es que los símbolos pueden ser latentes o despiertos (activados). Símbolo activo es aquel que ha sido desencadenado: es decir, aquel donde un número de neuronas suficiente para sobrepasar el umbral han sido excitadas por estímulos provenientes de fuera. Puesto que un símbolo puede ser desencadenado de muy diferentes formas, puede también actuar de muy diferentes formas, una vez despertado. Esto indica que debemos considerar el símbolo no como entidad fija, sino variable. Por consiguiente, para describir un estado cerebral no bastaría con decir “Los símbolos A, B, …, N están plenamente activos”; por el contrario, habría que incorporar un conjunto adicional de parámetros para cada símbolo activo, que caracterice determinados aspectos de las acciones que se cumplen internamente en los símbolos. Una pregunta muy interesante sería la de si existirán ciertas neuronas nucleares en cada símbolo, que se exciten invariablemente cada vez que el símbolo se activa; si tal conjunto nuclear de neuronas existe, podríamos referimos a él llamándolo el “núcleo invariante” del símbolo. Si siente la tentación de suponer que cada vez que se piensa en, digamos, una cascada, se repite el mismo proceso neural fijo, connotado de diversas maneras según el contexto, sin duda, pero volviendo a producirse sin excepción. No obstante, no es seguro que esto tenga que ser así.
Ahora bien, ¿qué hace un símbolo, una vez despertado? Una descripción de bajo nivel diría: “Se excitan muchas de sus neuronas”. Pero esto ya no nos interesa. La descripción de alto nivel eliminará toda referencia a las neuronas y se concentrará exclusivamente en los símbolos. Luego, una descripción de alto nivel de lo que hace un símbolo activo, a diferencia de un símbolo latente, diría, “Envía mensajes, o señales, cuyo objetivo es tratar de despertar, o desencadenar, otros símbolos”. Sin duda, estos mensajes son trasladados, bajo la forma de flujos de impulsos nerviosos, por neuronas; sin embargo, en la medida en que podamos evitar tal fraseología, debemos hacerlo, pues representa una perspectiva de bajo nivel de observación de los fenómenos, y esperar que nos sea posible actuar exclusivamente en un nivel alto. En otros términos, esperamos que sea posible tabicar los procesos del pensamiento, para su consideración, con respecto a los hechos neurales, del mismo modo que el comportamiento de un reloj es tabicado con respecto a las leyes de la mecánica cuántica o que la biología celular es tabicada con respecto a las leyes de los quarks.
¿Y cuál es la ventaja de esta representación de alto nivel? ¿Por qué es mejor decir “Los símbolos A y B desencadenaron el símbolo C”, que decir “Las neuronas 183 a la 612 estimularon la neurona 75 y causaron su excitación”? Esta interrogación fue respondida en la Furmiga: es mejor porque los símbolos simbolizan cosas y las neuronas no. Los símbolos son la concreción del hardware de los conceptos. Un grupo de neuronas que desencadena otra neurona es algo que no se corresponde con ningún acontecimiento exterior, en tanto que el desencadenamiento de cierto símbolo por otros símbolos guarda una relación con hechos del mundo real o de un mundo imaginario. Los símbolos se vinculan entre sí mediante los mensajes que envían de una parte a otra, en tal forma que sus patrones de desencadenamiento son muy semejantes a los acontecimientos de gran escala que suceden en nuestro mundo, o que podrían suceder en un mundo similar al nuestro. En esencia, la significación surge aquí por la misma razón que en el sistema mg: la presencia isomórfica; solamente que aquí el isomorfismo es infinitamente más complejo, sutil, delicado, versátil e intensional.
Dicho al margen, el requisito de que los símbolos sean capaces de trasladar mensajes complicados de una parte a otra quizá sea suficiente para excluir, en las neuronas, la posibilidad de que desempeñen el papel de símbolos. Como una neurona tiene exclusivamente una vía única de envío de la información desde sí misma, y carece de medios para dirigir selectivamente una señal en una o en otra dirección, no tiene, lisa y llanamente, el género de facultad selectiva de desencadenamiento del cual debe estar dotado un símbolo para actuar como un objeto del mundo real. En su libro The Insect Societies, E. O. Wilson hace una observación similar con respecto al modo en que se difunden los mensajes en el interior de una colonia de hormigas:
[La comunicación masiva] se define como la transmisión de información, dentro de grupos, que un individuo en particular no podría transmitir a otro.[3]
¡No es una imagen tan descartable: el cerebro, visto como una colonia de hormigas!
El problema que sigue —y es también de extrema importancia— se refiere a la naturaleza y a la “dimensión” de los conceptos representados, en el cerebro, por símbolos individuales. Las preguntas que se plantean acerca de la naturaleza de los símbolos son de este tipo: ¿Habrá un símbolo para la noción genérica de cascadas, o distintos símbolos para las diversas cascadas específicas? ¿O tendrán vigencia ambas variantes? En cuanto a la “dimensión” de los símbolos, los interrogantes serían: ¿Habrá un símbolo para cada narración completa? ¿Para cada melodía? ¿Para cada chiste? ¿O lo más probable sería que haya símbolos sólo para conceptos que tengan aproximadamente la dimensión de las palabras, y que las ideas más extensas, como las oraciones o ciertas expresiones, estén representadas por una activación simultánea o sucesiva de diversos símbolos?
Consideremos el tema de la dimensión de los conceptos representados por símbolos. La mayoría de los pensamientos están compuestos por elementos básicos, casi atómicos, a los que por lo general no sometemos a revisión. Su dimensión coincide, más o menos, con la de la palabra; a veces es algo mayor, a veces algo menor: por ejemplo, el nombre “catarata”, el nombre propio “Cataratas del Iguazú”, la desinencia “-aba” del pretérito imperfecto, la frase verbal “echar a perder”, y frases idiomáticas más extensas, están muy cerca de lo atómico. Son los trazos elementales de pincel que habitualmente usamos para pintar el retrato de conceptos más complejos, tales como el argumento de una película, la personalidad de una ciudad, la naturaleza de la conciencia, etc. Estas ideas complejas no son pinceladas individuales. Parece razonable pensar que las pinceladas de lenguaje son también pinceladas de pensamiento y que, por consiguiente, los símbolos representan conceptos de, aproximadamente, su misma dimensión. Así, un símbolo sería, de un modo general, algo que uno representa mediante una palabra o una expresión determinadas, o a lo cual es asociado un nombre propio. Y la representación cerebral de una idea más compleja, como por ejemplo un conflicto amoroso, sería una secuencia sumamente complicada de activaciones de diversos símbolos por la acción de otros símbolos.
Existe una distinción corriente con respecto al pensamiento: la que diferencia entre categorías e individuos, o entre clases y casos (otros dos términos utilizados en ocasiones son “tipos” y “especímenes”). Se podría suponer, a primera vista, que un símbolo dado tendría que ser, o bien el símbolo de una clase, o bien el símbolo de un caso: pero ello sería incurrir en una simplificación excesiva. En realidad, la mayoría de los símbolos pueden llenar ambos papeles, según el contexto de su activación. Por ejemplo, veamos esta lista:
(1) una publicación
(2) un periódico
(3) El País
(4) la edición del 5 de marzo de El País
(5) mi ejemplar de la edición del 5 de marzo de El País
(6) mi ejemplar de la edición del 5 de marzo de El País en el momento en que lo compré (en contraste con el mismo ejemplar días después, cuando lo uso para armar un barquito de papel)
Las líneas 2 a la 5 cumplen las dos funciones. Así, la línea 4 es un caso dentro de la clase general ubicada en la línea 3, y la línea 5 es un caso de la línea 4. La línea 6, por su parte, es un género especial de caso de una clase: una manifestación. Las manifestaciones de un objeto son los sucesivos estadios de éste en el transcurso de su historia vital. Es interesante preguntarse si las vacas de una granja perciben la identidad individual por debajo de todas las manifestaciones que se registran en el jovial granjero que las alimenta diariamente.
La lista anterior impresiona como una jerarquía en materia de generalidad, donde encontramos, arriba de todo, una muy amplia categoría conceptual, y en el último escalón una muy humilde cosa particularizada, y situada además en el espacio y en el tiempo. Sin embargo, la idea de que una “clase” debe ser siempre enormemente amplia y abstracta es demasiado limitada. Ello es así porque nuestro pensamiento se vale de un ingenioso principio, al que podemos denominar el principio del prototipo:
Aun el hecho más específico puede servir como ejemplo genérico de una clase de hechos.
Todos sabemos que los hechos específicos están dotados de una vividez que los imprime tan vigorosamente en la memoria que, más tarde, pueden ser utilizados como modelos de otros hechos, en alguna forma semejantes a aquéllos. Luego, en cada hecho específico existe el germen de una clase completa de hechos similares. Esta idea de existencia de generalidad en lo específico tiene alcances muy importantes.
Ahora bien, es natural que se formule la pregunta: ¿Los símbolos, en el cerebro, representan clases o casos? Y también éstas: ¿Hay ciertos símbolos que sólo representan clases, mientras que otros representan nada más que casos? ¿O bien un mismo símbolo puede actuar como símbolo de una clase, o como símbolo de un caso, según aquellas de sus partes que hayan sido activadas? Esta última presunción suena atractiva; se puede pensar que una activación “liviana” de un símbolo representaría una clase, y que una activación más profunda, o más compleja, contendría modelos más detallados de excitación neural interna, y en consecuencia representaría un caso. A la luz de una segunda reflexión, no obstante, ello resulta caprichoso: implicaría, por ejemplo, que la activación adecuadamente compleja del símbolo que representa a “publicación” derivaría en la obtención del muy complejo símbolo que representa un periódico específico cuyas páginas estoy empleando para hacer un barquito de papel. Y cualquier otra manifestación de toda otra porción de material impreso estaría internamente representada por alguna manera de activación del símbolo único que representa a “publicación”. Esto impresiona como una carga desmesuradamente pesada para poner encima del solo símbolo “publicación”. Es necesario concluir, por ende, que los símbolos de casos pueden coexistir, unos junto a los otros, con los símbolos de clases, y no que los primeros son exclusivamente modalidades de activación de los segundos.
Por otra parte, los símbolos de casos suelen heredar muchas de sus propiedades de las clases a las cuales pertenecen los casos. Si yo digo que fui a ver una película, mi interlocutor comenzará a “acuñar”, en ese momento, un nuevo símbolo de caso para esa película en particular; empero, si no aporto más información, el nuevo símbolo de caso tendrá que reposar en gran parte sobre el símbolo de clase representativo de “película”, preexistente en mi interlocutor. Inconscientemente, éste se atendrá a una multitud de presuposiciones acerca de la película: por ejemplo, la de que duró entre una y tres horas, la de que fue exhibida en una sala de esta misma ciudad, la de que narraba un relato sobre ciertas personas, y así sucesivamente. Estas presuposiciones forman parte de los símbolos de clase y constituyen los vínculos que se prevén con otros símbolos (es decir, relaciones de desencadenamiento potencial); se las puede denominar opciones subsidiarias. En todo símbolo de caso recién acuñado, las opciones subsidiarías pueden, sin dificultad, ser pasadas por alto; pero si aquél no es explícitamente formulado, permanecerán en él las opciones, recibidas en herencia de su símbolo de clase. Mientras no son descartadas, éstas suministran algunas bases preliminares para pensar en el nuevo caso —digamos, volviendo a nuestro ejemplo, para que mi interlocutor piense en la película que fui a ver—, mediante la utilización de conjeturas razonables, las cuales son aportadas por el “estereotipo” o símbolo de clase.
Un caso reciente y simple es como un niño sin ideas o experiencias propias: depende por completo de las experiencias y opiniones de sus padres, las cuales se limita a repetir mecánicamente. En forma gradual, sin embargo, a medida que interactúa cada vez más con el resto del mundo, el niño adquiere sus propias experiencias idiosincráticas, e inevitablemente comienza a apartarse de las de sus padres. Por último, el niño se convierte por completo en adulto. Del mismo modo, un caso nuevo puede separarse de su clase paterna luego de cierto lapso y llegar a ser una clase, o prototipo, por derecho propio.
A fin de ilustrar gráficamente este proceso de separación, supongamos que durante un viaje en automóvil sintonizamos una estación de radio que transmite un partido de fútbol entre equipos que no nos son familiares. Al principio, los nombres de los distintos jugadores nos resultan desconocidos. Cuando el locutor dice: “Remates es derribado en la zona del círculo central, cuando se disponía a cambiar de frente para el contraataque”, todo lo que registramos es que un jugador detuvo ilícitamente a otro. Ésta es una situación de activación del símbolo de clase “jugador de fútbol”, representativo de jugada violenta. Pero luego, cuando Remates aparece en algunas otras jugadas importantes, comenzamos a construir un nuevo símbolo de caso para él en particular, utilizando su nombre, quizá, como punto focal. Este símbolo es, igual que un niño, dependiente; está supeditado al símbolo de clase representativo de “jugador de fútbol”: casi toda nuestra imagen de Remates es suministrada por nuestro estereotipo de un jugador de fútbol, tal como está contenido en el símbolo “jugador de fútbol”. Paulatinamente, sin embargo, a medida que acumulamos mayor información, el símbolo “Remates” se hace más autónomo, y reposa cada vez menos en la activación concurrente de su símbolo paterno de clase. Esto puede ocurrir en el transcurso de escasos minutos, si Remates realiza algunas jugadas brillantes, que lo hagan destacarse. Sus compañeros de equipo, no obstante, pueden seguir siendo enteramente representados por activaciones del símbolo de clase. Por último, a lo mejor a los pocos días, cuando ya hemos leído determinados artículos en la sección deportiva del periódico, se rompe el cordón umbilical y Remates puede erguirse sobre sus propios pies: ahora sabemos cosas tales como cuál es su lugar de origen y dónde comenzó a jugar fútbol, reconocemos su rostro, etc. En este momento, Remates deja de ser concebido meramente como un jugador de fútbol y pasa a serlo como un ser humano quien, entre otras características, presenta la de jugar profesionalmente al fútbol. “Remates” es un símbolo de caso que puede transformarse en activo al tiempo que su símbolo paterno de clase (jugador de fútbol) permanece latente.
Al principio, el símbolo Remates fue un satélite que giraba alrededor de su símbolo madre, igual que un satélite artificial en vuelo alrededor de la Tierra, la cual es enormemente más grande y de mayor masa. Viene luego un estadio intermedio, donde uno de los símbolos tiene más importancia que el otro, pero puede considerarse que cada uno de los dos gira en tomo del restante, de modo semejante a como lo hacen la Tierra y la Luna. Finalmente, los nuevos símbolos se hacen autónomos y, a su vez, pueden actuar perfectamente como símbolos de clase alrededor de los cuales es posible que comiencen a rotar nuevos satélites: símbolos representativos de otras personas menos conocidas que Remates pero que tienen algo en común con él, por lo cual éste puede servirles transitoriamente de estereotipo, hasta tanto contemos con más información que permita a los nuevos símbolos, a su vez, adquirir autonomía.
Estos estadios de crecimiento y separación final de un caso con respecto a una clase se distinguirán entre sí de acuerdo a la manera en que se vinculan los símbolos abarcados. Indudablemente, a veces será muy difícil decidir con precisión dónde termina un símbolo y comienza el otro. ¿Cuán “activo” es un símbolo, en comparación con el otro? Si uno puede ser activado independientemente del otro, será entonces enteramente razonable considerarlo autónomo.
Con respecto a la metáfora astronómica que aplicamos hace un momento, es interesante señalar que el problema del movimiento planetario es extremadamente complejo; en realidad, el problema general que plantean tres cuerpos en interacción gravitacional (tales como la Tierra, la Luna y el Sol) está muy lejos de haber sido resuelto, aun después de varios siglos de estudio. Una situación en la cual es posible obtener soluciones muy aproximadas, no obstante, se presenta cuando uno de los cuerpos tiene mucha más masa que los otros dos (el Sol, en el caso citado); tiene sentido entonces considerar como estacionario a ese cuerpo, con los otros dos girando alrededor suyo; tras esto, finalmente, puede incluirse la interacción entre los dos satélites. Pero esta aproximación depende de que se fragmente el sistema: por un lado el Sol, y por otro una “agrupación”, es decir, el sistema Tierra-Luna. Esto es una aproximación, pero permite una comprensión sumamente profunda del sistema. Luego, ¿en qué medida dicha agrupación es una parte de la realidad, y en qué medida se trata de una elaboración mental, de una imposición humana de estructurar al universo? Este problema de la “realidad” de los límites trazados entre las agrupaciones que son percibidas como autónomas o semiautónomas generará un sinfín de dificultades cuando lo afrontemos con relación a los símbolos del cerebro.
Una cuestión de lo más desconcertante es la que plantea el simple tema de los plurales. ¿Cómo visualizamos, por ejemplo, tres perros en una taza de té? ¿O varías personas dentro de un elevador? ¿Empezamos con el símbolo de clase que representa a “perro”, al cual frotamos para obtener tres “ejemplares”? ¿O bien lo que hacemos es activar conjuntamente los símbolos “tres” y “perro”? La escena imaginada tendrá que incluir detalles, o de lo contrarío se hará difícil seguir sustentando la teoría. Por ejemplo, carecemos, por cierto, de un símbolo de caso independiente para cada nariz, bigote o rasgo que hayamos visto. Dejamos que los símbolos de clase se ocupen de esta gran cantidad de aspectos y cuando nos cruzamos en la calle con gente que tiene bigotes, simplemente activamos, de alguna manera, el símbolo de clase “bigote”, sin acuñar nuevos símbolos de caso, salvo que examinemos a esa gente en forma muy atenta.
Por otra parte, una vez que comenzamos a distinguir individuos ya no podemos apoyarnos en un solo símbolo de clase (“persona”, por ejemplo), y distribuirlo entre todos los diferentes seres que conocemos. Sin duda, deberán adquirir existencia separada los símbolos de caso representativos de dichos seres. Es ridículo suponer que esta hazaña pueda ser realizada mediante “malabarismos”, es decir, mediante el desplazamiento versátil de un mismo símbolo de clase entre diferentes modos de activación (uno para cada persona).
Tiene que haber lugar, entre los extremos, para muchos géneros de variantes intermedias. Puede que haya toda una jerarquía de modalidades de diferenciación, en el cerebro, entre símbolos de clase y símbolos de caso, que dé origen a símbolos —y a organizaciones de símbolos— dotados de distintos grados de especificidad. Los diversos tipos de activación individual y conjunta de símbolos que aparecen a continuación pueden ser responsables de imágenes mentales dotadas de grados diferentes de especificidad:
Esto nos devuelve de inmediato a la pregunta acerca de cuándo un símbolo se constituye en un nítido subsistema del cerebro. Consideremos, por ejemplo, la segunda instancia de la lista anterior: la activación simultánea de varios símbolos de clase, a través de alguna forma coordinada. Esto puede muy bien ser lo que ocurre cuando el concepto que consideramos es “sonata para piano” (donde los símbolos de “sonata” y de “piano” son, como mínimo, dos de los símbolos activados). Pero si este par de símbolos es activado en conjunto lo suficientemente a menudo, es razonable suponer que el vínculo entre ambos llegará a tener el vigor necesario para que funcionen como una unidad cuando sean activados juntamente, en la forma adecuada. Así, dos o más símbolos pueden actuar como si fuesen uno, bajo las condiciones debidas, lo cual significa que el problema de contar la cantidad de símbolos del cerebro es más laborioso de lo que uno podría sospechar.
En oportunidades, pueden surgir condiciones tales que dos símbolos anteriormente desvinculados sean activados en forma simultánea y a través de un modo coordinado. Es posible que se combinen tan bien entre sí que la suya impresione como una unión inevitable y surja de allí un nuevo símbolo, formado por la estrecha interacción de dos símbolos ya existentes. Si así ocurre, ¿habría razón para decir que el nuevo símbolo “siempre estuvo allí pero nunca había sido activado”, o habrá que afirmar que el nuevo símbolo ha sido “creado”?
Si esto parece excesivamente abstracto, apelemos a un ejemplo concreto: el Diálogo Canon Cangrejo. En la invención de este Diálogo, dos símbolos ya existentes —el de “canon cangrejo musical”, y el de “diálogo verbal”— tuvieron que ser activados simultáneamente y, en alguna medida, obligados a interactuar. Una vez hecho esto, el resto era completamente inevitable: nació un nuevo símbolo —un símbolo de clase— de la interacción de aquellos dos, el cual pudo, a partir de ese momento, ser activado por sí mismo. Ahora bien, ¿éste fue siempre un símbolo latente en mi cerebro? Si es así, también tiene que haber sido un símbolo latente en el cerebro de todo ser humano que siempre albergó los símbolos componentes, aun cuando en él no haya despertado nunca el nuevo símbolo. Esto querría decir que, para enumerar los símbolos dentro del cerebro de alguien, habría que incluir todos los símbolos latentes: todas las combinaciones y permutaciones posibles de todos los tipos de activaciones de todos los símbolos conocidos. Esto abarcaría también esas fantásticas criaturas de software que inventa nuestro cerebro mientras dormimos: esas extrañas mezclas de ideas que despiertan cuando su huésped se duerme… La existencia de tales “símbolos potenciales” muestra que es, realmente, una enorme simplificación el imaginar que el cerebro consiste en una colección bien definida de símbolos en estados bien definidos de activación. Captar un estado cerebral en función del nivel simbólico es algo mucho más arduo.
Ante el inmenso y siempre creciente repertorio de símbolos que tiene presencia en cada cerebro, cabe la pregunta de si finalmente no se llegará a un punto en que el cerebro se sature, esto es, en que ya no haya espacio para ningún símbolo más. Es posible que ello ocurriera si los símbolos jamás se superpusiesen entre sí, si ninguna neurona pudiese cumplir una doble función; entonces pasaría con los símbolos lo mismo que con las personas que ocupan un elevador. “¡Atención: este cerebro tiene una capacidad máxima de 350.275 símbolos!”.
Sin embargo, esto no constituye un rasgo necesario del modelo simbólico de la función cerebral. En rigor, la regla probablemente sea la superposición y el más extremo entrelazamiento de los símbolos, de modo tal que cada neurona, lejos de ser un miembro de un solo símbolo, probablemente sea parte funcional de centenares de símbolos. Esto conlleva una pequeña perturbación puesto que, si es cierto, ¿no ocurriría así que toda neurona sería parte de todo símbolo? En tal caso, no sería posible asignar una localización a los símbolos, porque cada uno de éstos estará identificado con el cerebro entero. Ello explicaría conclusiones como las que extrajo Lashley de sus extirpaciones de corteza en ratas… pero también significaría el abandono de nuestra idea original de fraccionar el cerebro en distintos subsistemas físicos. Nuestra primera caracterización de los símbolos “como concreciones del hardware de los conceptos” puede que sea, en el mejor de los casos, una tremenda simplificación. En verdad, si todo símbolo está construido con los mismos componentes que todo otro símbolo, carece de todo sentido hablar de símbolos diferentes. ¿Cuál sería la firma de la activación de un símbolo determinado, esto es, cómo se podría distinguir la activación del símbolo A de la activación del símbolo B? ¿Habrá que tirar toda nuestra teoría a la basura? Aun cuando no haya una superposición total de símbolos, ¿no será cada vez más difícil sostener nuestra teoría, en la medida en que la mayoría de los símbolos sí se superponen? (Una forma posible de representación de la superposición de los símbolos es presentada en la figura 68).
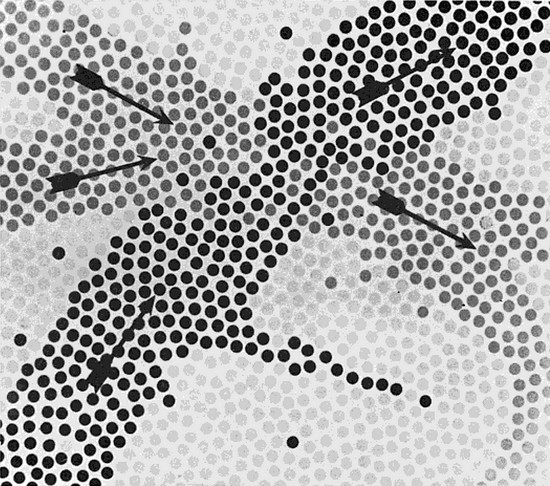
FIGURA 68. En este esquemático diagrama, las neuronas son imaginadas como puntos dispuestos en un plano. Se muestran dos recorridos neurales superpuestos, en diferentes matices de gris. Puede suceder que dos “destellos neurales” independientes, simultáneamente, transiten con la velocidad del rayo ambos recorridos, atravesándose como dos ondulaciones sobre la superficie de un lago (igual que en la figura 52). Esto ilustra la idea de dos “símbolos activos” que comparten neuronas y que, inclusive, pueden ser activados en forma simultánea. [De John C. Eccles, Facing Reality (New York: Springer Verlag, 1970), p. 21.]
Hay un medio para seguir sustentando una teoría basada en los símbolos, aun cuando éstos se superpongan, físicamente, de manera considerable o inclusive total. Consideremos la superficie de un lago, sobre la cual pueden producirse diferentes tipos de olas y ondulaciones. El hardware —a saber, el agua misma— es idéntico en todos los casos, pero cuenta con la posibilidad de distintos modos de excitación. Tales excitaciones, de nivel software, del mismo hardware, pueden ser diferenciadas perfectamente entre sí. No pretendo llevar tan lejos esta analogía como para sugerir que todos los símbolos son nada más que diferentes tipos de “ondulaciones” que se propagan a través de un medio neural uniforme, el cual no permite ninguna división significativa, en términos físicos, entre distintos símbolos. Pero puede suceder que, a fin de distinguir entre la activación de un símbolo y la activación de otro, sea necesaria la verificación de un proceso que involucre no solamente la localización de las neuronas que se excitan, sino también la muy precisa identificación de detalles relativos a la distribución temporal de las excitaciones que experimentan las neuronas. En otras palabras, ¿cuál neurona precede a cuál otra, y en qué medida?, ¿cuántas veces por segundo se excitó una neurona determinada? Quizá se vea así que pueden coexistir varios símbolos en el mismo conjunto de neuronas, gracias a características diferentes en cuanto a sus patrones de excitación neural. Entre una teoría que postule la distinción física de los símbolos, y otra que postule la superposición de símbolos, los cuales se distinguirían entre sí por sus modalidades de excitación, la diferencia consiste en que la primera aporta los elementos del hardware de los conceptos, en tanto que la segunda aporta elementos que corresponden parcialmente al hardware, parcialmente al software, de los conceptos.
De este modo, tenemos por delante dos problemas fundamentales para el desciframiento de los procesos del pensamiento, tal como éstos tienen lugar en el cerebro. Uno, es la necesidad de explicar cómo el tráfico de bajo nivel de las excitaciones neuronales da origen al tráfico de alto nivel de la activación de símbolos. El otro, la necesidad de explicar el tráfico de alto nivel de la activación simbólica de una manera intrínseca: elaborar una teoría que no aluda a los hechos neurales de bajo nivel. Si esto último es posible —y se trata de un supuesto clave, en el cual se apoyan todas las investigaciones actuales sobre inteligencias artificiales— la inteligencia, entonces, podrá estar concretada en tipos de hardware que no sean el cerebro. Así, se mostraría que la inteligencia es una propiedad que puede ser “desprendida” limpiamente del hardware en el cual reside; en otras palabras, que la inteligencia sería una propiedad software. Esto querrá decir que el fenómeno de la conciencia y de la inteligencia son, por cierto, de alto nivel, en el mismo sentido en que lo es la mayor parte de los restantes fenómenos complejos de la naturaleza: cuentan con sus propias leyes de alto nivel, las cuales dependen, aunque sean “desprendibles” de ellos, de los niveles más bajos. Si, por otro lado, no existe absolutamente ningún medio para concretar patrones de desencadenamiento de símbolos que carezcan por completo de un hardware de neuronas (o de neuronas simuladas), ello implicará que la inteligencia es un fenómeno ligado al cerebro, mucho más difícil de descifrar que otros cuya existencia se debe a una jerarquía de leyes vinculadas a la presencia de varios niveles diferentes.
En este punto nos volvemos a plantear el misterioso comportamiento colectivo de las colonias de hormigas, quienes pueden construir gigantescos e intrincados hormigueros, a pesar del hecho de que las, aproximadamente, 100.000 neuronas del cerebro de una hormiga no contienen, casi seguramente, ninguna información relativa a la estructura de un hormiguero. ¿Cómo es entonces que surge el hormiguero? ¿Dónde reside la información? En particular, cabe preguntarse dónde se encontrará la información que describe un arco como el que aparece en la figura 69. De alguna manera, ésta debe hallarse diseminada en la colonia, en la distribución de castas, en la distribución por edades y, probablemente, de modo muy amplio en las propiedades físicas del propio cuerpo de las hormigas. Es decir, la interacción entre hormigas está determinada en igual medida por la circunstancia de que cuentan con seis patas, cierto tamaño, etc., como por la información almacenada en su cerebro. ¿Podrá haber una Colonia Artificial de Hormigas?
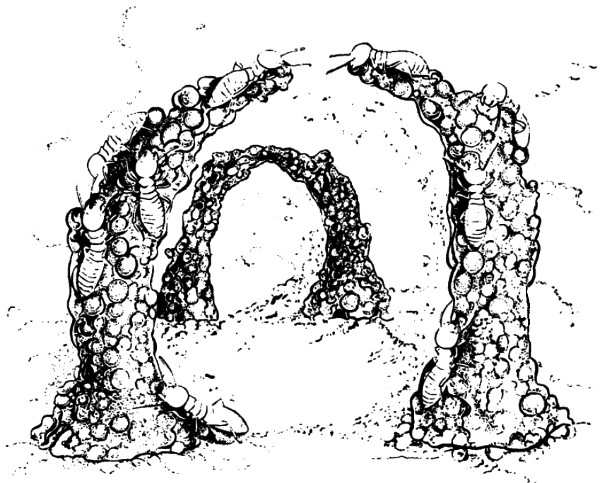
FIGURA 69. La construcción de un arco por trabajadoras de las termitas Macrotermes bellicosus. Cada columna es construida mediante la acumulación de bolitas de tierra y excremento. En la parte externa de la columna de la izquierda se observa una obrera en el momento en que deposita una bolita fecal. Otras, que han transportado bolitas en sus mandíbulas a lo largo de las columnas, las están colocando en los extremos en crecimiento de las mismas. Cuando una columna alcanza determinada altura, las termitas, guiadas evidentemente por el olor, comienzan a dirigirlas en ángulo, hacia la columna vecina. Al fondo, se ve un arco ya completado. [Dibujo de Turid Hölldobler; tomado de E. O. Wilson, The Insect Societies (Cambridge, Mass.: Harvard University Press, 1971), p. 230.]
¿Es posible que un único símbolo pueda ser despertado, en forma separada con respecto a todos los demás? Es probable que no. Del mismo modo que los objetos del mundo están siempre en un contexto formado por otros objetos, los símbolos siempre están conectados a una constelación de otros símbolos. Esto no significa, necesariamente, que los símbolos no puedan ser desentrelazados entre sí. Para utilizar una analogía bastante simple: en una especie, aparecen siempre a un tiempo especímenes masculinos y femeninos; sus funciones están completamente interrelacionadas y sin embargo ello no quiere decir que no pueda distinguirse entre un espécimen masculino y uno femenino. Cada uno es reflejado por el otro, igual que las cuentas de la red de Indra se reflejan entre sí. El entrelazamiento recursivo de la funciones F(n) y M(n), en el Capítulo V, no impide que cada función tenga sus características propias. El entrelazamiento de F y M podría ser representado por dos RTR que apelen una a la otra. Desde aquí podemos brincar a toda una red de RTA imbricadas entre sí: una heterarquía de procedimientos recursivos interactivos. En este campo, el entramado es tan inherente que ninguna RTA puede ser activada en forma separada; sin embargo, su activación puede ser totalmente peculiar, imposible de confundir con la de ninguna otra RTA. ¡No es una imagen tan descartable: el cerebro, visto como una colonia de RTA!
También los símbolos, a pesar de los múltiples lazos que los entremezclan, deben permitir que se los aísle. Esto podría implicar la identificación de una red neural, una red más una modalidad de excitación, o bien, probablemente, de algo perteneciente a un género completamente distinto. De todos modos, si los símbolos son parte de la realidad, es presumible que exista un procedimiento natural para trazar su diseño en un cerebro real. Aun así, en el caso de que finalmente fuesen identificados algunos símbolos en un cerebro, esto no querría decir que cualquiera de ellos pueda ser despertado en forma aislada.
El hecho de que un símbolo no pueda ser despertado en forma aislada no opaca la identidad independiente del símbolo; en realidad, se trata de lo contrario: la identidad de un símbolo se funda precisamente en su forma de conectarse (a través de los vínculos de desencadenamiento potencial) con el resto de los símbolos. La red mediante la cual los símbolos tienen la capacidad potencial de desencadenarse entre sí constituye el modelo que la actividad cerebral ofrece el universo real y de los universos alternativos que tome en consideración (los cuales son tan importantes para la supervivencia del individuo en el mundo real como lo es el mundo real mismo).
Nuestra facilidad para convertir clases en casos, y casos en clases, reside en las bases de nuestra inteligencia, y se trata de una de las grandes diferencias entre el pensamiento humano y los procesos de pensamiento de otros animales. No es que yo haya pertenecido a otras especies, y experimentado en carne propia cómo es pensar a la manera de éstas, sino que, desde fuera, es evidente que ninguna otra especie forma conceptos generales como lo hacemos nosotros, o imagina mundos hipotéticos: variantes del mundo que conocemos, las cuales ayudan a resolver qué trayectorias futuras elegir. Veamos, por ejemplo, el famoso “lenguaje de las abejas”, que consiste en danzas dotadas de información, ejecutadas por las obreras, de retorno a la colmena, para comunicar al resto de las abejas la ubicación de los sitios ricos en néctar. Aunque haya en cada abeja un conjunto de símbolos rudimentarios, que son activados por dicha danza, no hay motivo para creer que una abeja cuente con un vocabulario ampliable de símbolos. Las abejas y otros insectos parecen no tener la facultad de generalizar, esto es, de desarrollar nuevos símbolos de clase a partir de casos que son percibidos como casi idénticos.
Un experimento clásico es el realizado con avispas solitarias, el cual es narrado en el libro Mechanical Man, de Dean Wooldridge, al que pertenece este pasaje:
Cuando llega la época de depositar sus huevos, la avispa Sphex construye para ello una cueva, y se procura un grillo, al que pica de modo, no de matarlo, sino de paralizarlo. Lo arrastra luego al interior de la cueva, deposita allí sus huevos, la cierra y levanta vuelo para no retomar jamás. A su debido tiempo, la cría rompe el cascarón y se alimenta del paralizado grillo, el cual no ha sufrido descomposición alguna pues ha sido conservado en el equivalente avispa de una congelación a muy baja temperatura. Para el pensamiento humano, un procedimiento tan elaboradamente organizado y a todas luces dotado de finalidad impresiona como guiado por la lógica y la reflexión… antes del examen de otros detalles. Por ejemplo, la rutina de la avispa consiste en llevar al paralizado grillo hasta la cueva, dejarlo en el umbral, ingresar para asegurarse de que todo está en orden, salir y por último arrastrar al grillo dentro de la cueva. Si el grillo se aparta algunos centímetros en el transcurso de la inspección mencionada, la avispa, al salir de la cueva, volverá a poner al grillo en el umbral, no en el interior, y repetirá el procedimiento preparatorio de ingresar a la cueva para verificar que todo esté como corresponde. Si de nuevo el grillo se desplaza unos centímetros mientras la avispa está adentro, ésta volverá a ubicarlo en el umbral y otra vez ingresará a la cueva para efectuar el control final. La avispa nunca considera la posibilidad de llevar, en estos casos, al grillo directamente al interior de la cueva. En cierta oportunidad, este procedimiento fue repetido cuarenta veces, siempre con el mismo resultado.[4]
Da la impresión de que este comportamiento cuenta con un hardware completo. Pueden existir, en el cerebro de la avispa, símbolos rudimentarios, capaces de desencadenarse entre sí, pero no algo semejante a la capacidad humana de concebir distintos casos como casos de una clase aún-no-conformada y, en consecuencia, elaborar el símbolo de clase; tampoco existe en el cerebro de la avispa algo semejante a la capacidad humana de preguntarse, “¿Qué ocurre si hago tal cosa, qué resultados tendría en ese mundo hipotético?”. Este tipo de pensamiento requiere la capacidad de construir casos y operarlos como si fuesen símbolos que representan objetos comprendidos por una situación real, aunque tal situación no se esté produciendo e incluso no se produzca jamás.
Volvamos a la broma de Inocentes acerca del automóvil prestado y las imágenes suscitadas en nosotros durante la comunicación telefónica. Para la formación de éstas se necesita, primeramente, que se activen los símbolos representativos de un camino, de un automóvil, de una persona dentro de un automóvil. Ahora bien, el concepto “camino” es muy general y quizás incluya un repertorio de variantes que, de modo inconsciente, podemos extraer de la memoria latente cuando es oportuno hacerlo. “Camino” es una clase, antes que un caso. Cuando escuchamos el relato, rápidamente activamos símbolos que son casos revestidos de una especificidad en incremento gradual. Por ejemplo, cuando nos enteramos de que el camino estaba húmedo, ello suscita una imagen más específica, pese a nuestra comprensión de que, muy probablemente, dicha imagen difiera por completo del camino verdadero donde ocurrió el accidente. Pero esto no es importante: lo que interesa es si nuestros símbolos son lo debidamente adecuados como para seguir el relato, es decir, si los símbolos que el relato desencadena pertenecen al género correcto.
A medida que avanza la narración, completamos más aspectos de este camino: hay un borde elevado contra el cual un automóvil podría destrozarse. Ahora bien, ¿esto significa que estamos activando el símbolo de “borde” o que estamos fijándole algunos parámetros a nuestro símbolo de “camino”? Indudablemente, ambas cosas. Es decir, la red de neuronas que representa a “camino” tiene diferentes vías de excitación; lo que estamos haciendo es seleccionar la subred a la que toca excitarse en este instante. Al mismo tiempo, estamos activando el símbolo de “borde”, lo cual tiene, probablemente, una función instrumental en el proceso de selección de los parámetros de “camino” ya que las neuronas de “borde” pueden enviar señales a algunas de las involucradas en “camino” y viceversa. (Si esto suena un tanto confuso, es porque, en alguna medida, no estoy distinguiendo lo suficiente entre niveles de descripción: estoy tratando de elaborar una representación de los símbolos y simultáneamente de las neuronas que lo componen). Los verbos y las preposiciones no son menos importantes que los sustantivos. También ellos activan símbolos, los cuales se dirigen mensajes entre sí, en todos los sentidos. Por supuesto, hay diferencias características entre los patrones de desencadenamiento de símbolos correspondientes a los verbos y los correspondientes a los sustantivos, lo cual significa que pueden estar organizados, físicamente, de un modo algo distinto. Por ejemplo, puede que los sustantivos cuenten con símbolos perfectamente localizados, mientras que los verbos y las preposiciones quizá tengan símbolos dotados de muchos “tentáculos” diseminados por toda la corteza; o bien, una cantidad enorme de otro tipo de posibilidades.
Luego de completada la narración, se nos hace saber que no es verdadera. La facultad de obtener casos “por frotación”, a partir de las clases, como quien saca brillo a la broncería de un templo, nos ha permitido representarnos la situación y nos ha liberado de la necesidad de permanecer fieles al mundo real. El hecho de que los símbolos puedan actuar como matrices de otros símbolos nos brinda cierta independencia mental con respecto a la realidad: podemos crear universos artificiales, donde sucedan acontecimientos ficticios caracterizados por todos los detalles que se nos ocurra asignarles. Sin embargo, los símbolos de clase mismos, de los cuales emana toda esta prodigalidad, están profundamente enraizados en la realidad.
Por lo común, los símbolos cumplen funciones isomórficas con respecto a acontecimientos que verosímilmente puedan suceder, aunque a veces sean activados símbolos que representan situaciones que no pueden ocurrir, como la de una tuba poniendo huevos, por ejemplo, o la de dos torres de ajedrez caminando del brazo. Los límites entre lo que puede suceder y lo que no puede suceder son extremadamente difíciles de trazar. Cuando imaginamos un hecho hipotético, ponemos a ciertos símbolos en estado activo: según cómo interactúen (lo cual se reflejará, presumiblemente, en nuestro grado de asentimiento a continuar en la misma línea de pensamiento), es que diremos si el hecho “puede” o “no puede” suceder. De modo, pues, que las expresiones “puede” y “no puede” son sumamente subjetivas. En realidad, hay un amplio acuerdo entre la gente con respecto a cuáles acontecimientos pueden o no pueden ocurrir. Ello muestra el gran volumen de estructura mental que compartimos por entero… pero hay una área fronteriza donde se hace evidente el carácter subjetivo de la decisión acerca de qué mundos hipotéticos estamos dispuestos a tener en cuenta. Un estudio cuidadoso de los géneros de hechos imaginarios que los seres humanos estiman que pueden y no pueden producirse haría posible profundizar nuestro conocimiento de los patrones desencadenantes de símbolos, gracias a los cuales pensamos.
Cuando finaliza el relato, tenemos construido por completo un elaborado modelo mental de la escena, y en este modelo todos los objetos obedecen las leyes físicas. Ello quiere decir que las leyes físicas deben estar presentes, implícitamente, en los patrones desencadenantes de los símbolos. La expresión “leyes de la física” no significa aquí, por cierto, “las leyes de la física tal como son expuestas por los físicos”, sino las leyes intuitivas y articuladas en bloques que todos necesitamos tener en nuestras mentes a fin de sobrevivir.
Es curioso que podamos, voluntariamente, elaborar secuencias mentales de hechos que violan las leyes físicas, si así lo deseamos. Por ejemplo, si sugiero a alguien que imagine una escena donde dos automóviles se aproximan de frente y luego pasan uno a través del otro, esa persona no tendrá dificultades para conseguirlo. Las leyes físicas intuitivas pueden ser desplazadas por las leyes imaginarias de la física; ahora bien, cómo se produce este desplazamiento, cómo son elaboradas tales secuencias de imágenes y, ciertamente, qué clase de imágenes visuales son éstas, son misterios profundamente escondidos: inaccesibles piezas de conocimiento.
Es innecesario recordar que, en nuestro cerebro, no solamente hay leyes articuladas en bloques, relativas al modo en que actúan los objetos inanimados, sino también al modo en que lo hacen las plantas, los animales, la gente y las sociedades; en otras palabras, leyes articuladas en bloques de la biología, la psicología, la sociología, y así sucesivamente. Todas las representaciones internas de tales entidades implican la presencia inevitable de modelos construidos en función de bloques: el determinismo es sacrificado a la simplicidad. Nuestra representación de la realidad no va más allá de poder predecir las probabilidades de alcanzar determinadas zonas, dentro de espacios abstractos de comportamiento: no puede predecir nada con la precisión de la física.
En el campo de las inteligencias artificiales se distingue entre conocimiento de tipo procedimental y de tipo declarativo. Se dice de una pieza de conocimiento que es declarativa cuando es explícitamente almacenada, de modo que no solamente el programador, sino también el programa, puedan “leerla” como si estuviera incluida en una enciclopedia o en un repertorio. Esto quiere decir, generalmente, que su codificación está localizada y no ampliamente diseminada. En contraste, el conocimiento procedimental no se codifica como hecho, sino como programa. Un programador puede estar en condiciones de hacer un examen y luego decir, “Veo que gracias a estos procedimientos incluidos aquí, el programa ‘sabe’ escribir oraciones en inglés”, pero el programa mismo puede no ser explícitamente sabedor de cómo hace para escribir esas oraciones. Por ejemplo, su vocabulario quizá no incluya, para nada, las palabras inglesas que significan “inglés”, “oración” y “escribir”… Luego, el conocimiento procedimental está diseminado por todas las piezas, y no se lo puede delimitar, ni “teclear”: es una consecuencia global del modo en que funciona el programa, no un detalle localizado. En otras palabras, una pieza de conocimiento exclusivamente procedimental es un epifenómeno.
En la mayoría de las personas coexiste, junto con una vigorosa representación procedimental de la gramática de su lengua materna, una representación declarativa, más débil, de la misma. Ambas pueden entrar fácilmente en conflicto, como se advierte cuando un hablante nativo enseña a un extranjero, a menudo, a utilizar expresiones que él mismo no emplearía nunca, pero que emanan del declarativo “libro de texto” que conoció en su época escolar. Las leyes intuitivas o articuladas en bloques de la física y de las demás disciplinas ya mencionadas se ubican principalmente en el sector procedimental; el conocimiento de que un pulpo tiene ocho tentáculos se ubica principalmente en el sector declarativo.
Entre los extremos declarativo y procedimental hay todos los matices posibles. Consideremos el recuerdo de una melodía: ¿se encuentra ésta almacenada en nuestro cerebro, nota por nota? ¿Un cirujano podría extraer un rollo de filamento neural de nuestro cerebro, estirarlo y luego proceder a señalar a lo largo del mismo las notas sucesivamente almacenadas, casi como si se tratara de una porción de cinta magnética? Si es así, las melodías están almacenadas declarativamente. ¿O bien el recuerdo de una melodía pasa por la mediación de la interacción de un gran número de símbolos, algunos de los cuales representan relaciones tonales; otros, cualidades emotivas, otros, aspectos rítmicos, etc.? Si es así, el almacenamiento de las melodías es procedimental. En verdad, probablemente haya una mezcla de ambas variantes en el modo en que una melodía es almacenada y recordada.
Es interesante que cuando se extrae una melodía de la memoria la mayoría de las personas no discrimina en cuanto a la tonalidad, y así es como “Cumpleaños feliz” se canta tanto en fa sostenido como en do. Esto indica que son almacenadas las relaciones entre sonidos y no los sonidos absolutos. Pero no hay motivo para que una relación entre sonidos no pueda ser almacenada en forma totalmente declarativa. Por otro lado, algunas melodías son muy fáciles de memorizar, pero existen otras sumamente huidizas; si sólo fuera cuestión de acopiar notas sucesivas, sería igualmente fácil almacenar una melodía como cualquier otra. El hecho de que algunas melodías sean pegadizas y otras no parece indicar que el cerebro tiene cierto repertorio de patrones familiares, los cuales son activados cuando es escuchada la melodía. Así, para “ejecutar” la melodía, esos patrones tendrían que ser activados en el mismo orden, y esto nos lleva de nuevo a la noción de símbolos que se desencadenan entre sí, y no a la de una simple secuencia lineal de notas, o relaciones entre sonidos, almacenados declarativamente.
¿Cómo sabe el cerebro si una pieza de conocimiento está almacenada declarativamente? Por ejemplo, supongamos que se nos pregunta, “¿Cuántos habitantes tiene la ciudad de Barcelona?”: una cifra aproximada a los tres millones nos vendrá a la mente, y no una respuesta sorprendida del tipo, “Oiga, ¿cómo se le ocurre que voy a ponerme a contarlos?”. Supongamos ahora que la pregunta es, “¿cuántas sillas hay en la sala de su casa?”. Aquí sucede lo contrario: en lugar de rastrear la respuesta en un fichero mental, nos dirigimos de inmediato a la sala y contamos las sillas, o bien reconstruimos la sala en nuestra cabeza y contamos las sillas que aparezcan en esa imagen. Las dos preguntas anteriores son del mismo tipo —“¿cuántas?”—, pero una de ellas trajo como resultado la recuperación de una pieza de conocimiento declarativo, mientras que la otra trajo como resultado que se recurriera a un método procedimental de búsqueda. Está claro, según lo muestra este ejemplo, que tenemos conocimiento de cómo clasificamos nuestro propio conocimiento y, lo cual es más importante, que una parte de ese metaconocimiento puede, a su vez, ser almacenado de manera procedimental, de modo que sea utilizado sin que, inclusive, advirtamos cómo se produce tal utilización.
Una de las cualidades más notables y más difíciles de describir de la conciencia es la imaginación visual. ¿Cómo creamos una imagen visual de la sala de nuestra casa? ¿O de un rápido arroyo montañés? ¿O de una naranja? Más misteriosamente todavía: ¿cómo construimos imágenes inconscientemente, imágenes que guían nuestros pensamientos, a los cuales otorgan fuerza, color, profundidad? ¿De qué almacén son recuperadas? ¿Qué magia nos permite entremezclar dos o tres imágenes, sin brindar ningún indicio acerca de cómo lo conseguimos? El conocimiento de cómo hacemos esto se encuentra entre los más procedimentales de todos, pues no tenemos casi ningún vislumbre en cuanto a qué es la imaginación mental.
Puede que la imaginación se base en nuestra capacidad de suprimir la actividad motriz. Con esto, quiero decir lo siguiente: si imaginamos una naranja, en nuestra corteza puede suscitarse un conjunto de órdenes, tales como recogerla, olería, examinarla, etc. Sin duda, estas órdenes no pueden ser cumplidas porque la naranja no está allí, pero pueden ser dirigidas, a través de los canales ordinarios, hacia el cerebelo u otros subórganos del cerebro hasta que, en algún punto crítico, se cierre un determinado “grifo mental”, impidiendo así que las órdenes sean llevadas a la práctica. Según el grado de avance, en la línea donde se sitúe el “grifo”, las imágenes podrán ser más o menos vívidas y verosímiles. La cólera puede llevamos a imaginar con total verismo que tomamos un objeto y lo arrojamos, o que le damos una patada a alguna cosa; sin embargo, no procedemos efectivamente así. Por otra parte, nos sentimos “casi” actuando efectivamente así. Es probable que el grifo detenga los impulsos nerviosos “en el último instante”.
Hay otra vía a través de la cual la visualización hace notar la distinción entre conocimiento accesible e inaccesible. Examinemos cómo fue nuestra visualización de la escena del automóvil resbalando sobre el camino de montaña. Por supuesto, imaginamos las montañas mucho más grandes que el automóvil. Ahora bien, ¿esto ocurre porque en alguna oportunidad, hace mucho, tuvimos ocasión de advertir que “los automóviles no son tan grandes como las montañas”, fijamos entonces el dato en nuestra memoria, y al imaginarnos lo que nos relata nuestra amiga recuperamos dicho dato y lo utilizamos para construir nuestra imagen? Se trata de una teoría muy improbable. ¿Esto no ocurrirá, en cambio, como consecuencia de ciertas interacciones, introspectivamente inaccesibles, de los símbolos que han sido activados en nuestro cerebro? Obviamente, esto último parece mucho más verosímil. El conocimiento de que los automóviles son más pequeños que las montañas no es una pieza de memorización mecánica, sino una pieza de conocimiento que puede ser generada por deducción. En consecuencia, lo más probable es que no esté almacenada en ningún símbolo individual de nuestro cerebro sino que, por el contrario, pueda producirse como resultado de la activación, seguida de interacción recíproca, de muchos símbolos: los que representan a “comparar”, “tamaño”, “automóvil”, “montaña”, pongamos por caso, y probablemente algunos otros. Esto significa que el conocimiento no está acopiado explícita, sino implícitamente, a través de una muy amplia diseminación, y no bajo la forma de un “paquete de información” localizado. Circunstancias tan simples como el tamaño relativo de tos objetos tienen que ser reunidas, en lugar de, simplemente, recuperadas. Por consiguiente, inclusive en el caso de una pieza verbalmente accesible de conocimiento, existen complejos procesos inaccesibles que actúan como intermediarios para que se pueda llegar al punto en que aquélla esté lista para ser dicha.
Continuaremos con nuestra exploración de las entidades denominadas “símbolos” en diferentes capítulos. En los Capítulos XVIII y XIX, sobre inteligencias artificiales, consideraremos algunas formas posibles de instrumentar símbolos activos en programas. En el capítulo que sigue, veremos ciertos aportes, referidos a la comparación entre cerebros, que nos brinda nuestro modelo de base simbólica de la actividad cerebral.