
Nada mejor, para finalizar un libro como el presente, que ocuparse de un apartado de la ciencia que está protagonizando el tiempo en que vivimos, un tiempo dominado por una profunda revolución científica: la biología molecular.
Vivimos, en efecto, en una época científicamente revolucionaria. Se trata, además, de una revolución con características muy especiales: al contrario que otras revoluciones científicas de tiempos pasados —como la newtoniana, la lavoisieriana, la einsteiniana o la promovida por la física cuántica (la planckiana, podríamos acaso denominarla)—, esta tiene que ver de una forma mucho más directa con nosotros mismos, en tanto que tiene como objetivo el estudio de la vida, incluyendo, claro está, la vida humana. Mientras los temas, los protagonistas, de revoluciones tan recientes como la relativista y la cuántica se llamaban «espacio-tiempo», «átomos», «principio de incertidumbre», «fisión nuclear», «niveles energéticos», «transistores», «agujeros negros» o «universo en expansión», en la revolución biológica molecular en curso esos temas, esos protagonistas, se denominan «ADN», «genes», «transgénicos», «clonación» o «células madre», conceptos que evidentemente, en sentido inmediato, nos son mucho más cercanos.
Nos encontramos, en definitiva, justo en medio (al principio tal vez) de una revolución con profundas implicaciones no sólo en el dominio de la ciencia, sino también en el social. Estamos asistiendo, y siendo creadores al igual que objetivos, de uno de esos períodos en los que, por decirlo de alguna manera, el suelo tiembla bajo nuestros pies, en el que nos damos cuenta de que el futuro será diferente del presente y de que el propio presente cambia, debido precisamente a los instrumentos que se van creando dentro de esa revolución.
Los caminos de la vida: cromosomas y genes
Como ya señalé en el capítulo 4, a mediados del siglo XIX Gregor Mendel desarrolló la teoría según la cual ciertos elementos discretos, sobre cuya estructura él poco podía decir, son responsables de los rasgos hereditarios, un resultado al que llegó cruzando variedades diferentes de guisantes y observando los productos de tales hibridaciones en generaciones sucesivas. En el lenguaje actual, distinguió entre la apariencia de un organismo (su fenotipo), que, por supuesto, puede ser una mezcla de las características de sus progenitores, y el conjunto de factores (genes) heredados de cada uno de los padres (el genotipo), que continúa siendo discreto y sin mezcla.
Mendel (Museo de Moravia, Brno).
También mencioné que la esencia de las ideas de Mendel pasó desapercibida para la comunidad de biólogos hasta su redescubrimiento en 1900, gracias a los trabajos de Hugo de Vries, Carl Correns y Eric von Tschermak. Esto fue así porque difícilmente sus contemporáneos podrían haber relacionado el concepto de elementos hereditarios discretos con las nociones existentes entonces en el dominio de la biología celular, sin olvidar que la metodología estadística mediante la cual Mendel interpretó sus resultados era ajena al modo de pensamiento vigente en aquella época. Su artículo de 1865 fue redescubierto cuando ya se había introducido el concepto de cromosomas, la «primera aproximación a la unidad discreta de herencia».
Pasajes de «Versuche über Pflanzen-Hybriden» («Experimentos sobre la hibridación de plantas») de Mendel (1865):
Observaciones preliminares.
El tema de los ensayos de los que se hablará en este trabajo surgió de una serie de fecundaciones artificiales realizadas en plantas ornamentales con el fin de obtener nuevas variantes de color. La sorprendente regularidad con que reaparecían las mismas formas híbridas cada vez que se realizaba la fecundación entre las mismas especies, sugirió otros experimentos complementarios para investigar el desarrollo de los descendientes de esos híbridos.
Observadores tan minuciosos y concienzudos como Kölreuter, Gärtner, Herbert, Lococq, Wichura y otros han dedicado con incansable perseverancia una parte de su vida a ese problema. En particular, fue Gärtner quien, en su obra La obtención de mestizos en el reino vegetal, reunió observaciones altamente valiosas. Últimamente, Wichura ha publicado sus profundas investigaciones sobre los híbridos de los sauces. El que hasta ahora no haya sido posible formular una ley general sobre la formación y el desarrollo de los híbridos, no puede extrañar a nadie que, conociendo el alcance del problema, sepa apreciar las dificultades que hay que vencer en ensayos de este tipo. Sólo se podrá obtener una solución definitiva cuando se realicen experimentos detallados con las diversas familias de plantas. El que examine los trabajos existentes en esta materia llegará a la convicción de que ninguno de los numerosos experimentos conocidos ha sido realizado en la escala y en la forma necesarias para determinar las cantidades necesarias de las diferentes formas bajo las cuales aparecen los descendientes de los híbridos, para ordenar tales formas con seguridad en las distintas generaciones y determinar las relaciones numéricas respectivas…
El presente artículo trata de un intento de tal experimento de detalle. Según correspondía, se limitó a un pequeño grupo de plantas y está ahora, después de ocho años, prácticamente finalizado.
…
La configuración de los híbridos.
Experimentos realizados en años anteriores con plantas ornamentales ya demostraron que los híbridos no representan siempre un término medio entre las especies paternas. En algunos caracteres que saltan a la vista, como los que se refieren a la forma y el tamaño de las hojas, a la pubescencia de las distintas partes, etc., resulta evidente la estructura intermedia; en otros casos, en cambio, es tan grande la supremacía de uno de los dos caracteres paternos, que resulta difícil o completamente imposible descubrir el otro progenitor en el híbrido.
Ahora bien, no fue en modo alguno fácil llegar a una teoría de la reproducción y de la herencia basada en elementos discretos. De hecho, cuando se analiza la historia de la biología correspondiente a, básicamente, las décadas de 1870 y 1880, no es difícil detectar que los indudables avances que se fueron realizando coexistieron con planteamientos menos atinados. Y es que, seguramente la mayor parte de las veces la creación de nuevas teorías constituye un complejo y delicado proceso en el que lo viejo —los conceptos, expectativas o, simplemente, «modos de mirar la naturaleza» asumidos hasta entonces— se mezcla con lo nuevo, en una combinación en ocasiones explosiva de la que surge con dificultad, y no siempre rápidamente, una visión, un modelo, diferente.
Un ejemplo magnífico en este sentido es el del catedrático de zoología y anatomía comparada en la Universidad de Friburgo, August Weismann (1834-1914), considerado como uno de los fundadores de la genética moderna. Ya hacia 1868, Weismann llegó a la conclusión de que la «dirección de desarrollo» se transfería por medio del protoplasma del esperma y de la célula del huevo. A través de consideraciones evolucionistas e investigando la cuestión de si se podían heredar caracteres adquiridos, Weismann (que era un ferviente darwinista) desarrolló su teoría del «plasma germinal», basada en la existencia de una sustancia responsable de la transmisión de los rasgos hereditarios (él argumentaba —contra Lamarck— que los caracteres adquiridos no se pueden heredar, ya que en tal caso tendrían que afectar al propio plasma germinal). Estrictamente, no fue él el único en pensar en base a la continuidad de una «sustancia de la herencia». En 1872, por ejemplo, Francis Galton (1822-1911), más conocido por sus tesis eugenésicas, la había vislumbrado, pero es a Weismann a quien se le debe adjudicar el crédito de haber desarrollado esa teoría en un conjunto más o menos coherente, un conjunto que fue reelaborando para ponerlo en relación con los resultados de las nuevas investigaciones sobre la estructura celular. En 1883, todavía concebía las células germinales como portadoras de configuraciones de moléculas que conducían a las células reproductivas. Estaba, en definitiva, tratando aún con la cuestión de la herencia en términos de toda la célula (germinal), y es desde semejante perspectiva como se debe entender su manifestación (recogida en obras suyas como Das Keimplasma [El plasma germinal], de 1892, o en la edición inglesa de sus ensayos completos: Essays upon Heredity and Kindred Biological Subjects [Ensayos sobre herencia y temas biológicos relacionados], publicada en 1891) de que la reproducción ponía en juego células de un tipo particular, las células germinales (o sexuales), frente a las somáticas, o corporales, que contienen una sustancia, decía, «cuya constitución físico-química, incluida su naturaleza molecular, le otorga la facultad de convertirse en un nuevo individuo de la misma especie».
Sin embargo, y al hilo de los resultados que iban obteniendo otros investigadores, en los años siguientes su concepto del plasma germinal fue cambiando. En particular, las décadas de 1870 y 1880 constituyeron un período muy activo en hallazgos que terminaron imponiendo la idea de que es en el núcleo de las células donde tiene lugar el proceso que controla la reproducción (una idea esta que de alguna manera ya había defendido por razones teóricas el zoólogo germano Ernst Haeckel [1834-1919] en 1866). Un nombre que hay que resaltar en este sentido es el de Walther Flemming (1843-1905), catedrático de anatomía en la Universidad de Kiel, quien, a partir de investigaciones de las divisiones celulares en anfibios (describió y bautizó el proceso de la «mitosis»), reunidas en su texto Zellsubstantz, Kern und Zelltheilung (Sustancia celular, núcleo y división celular; 1882), fue capaz de, primero, diferenciar en los núcleos celulares una parte fuertemente teñible, la cromatina, y una parte muy poco teñible, la acromatina, y, después, poner de manifiesto que durante la división celular, los gránulos de cromatina se disponían en filamentos, a los que más tarde, en 1888, el anatomista de la Universidad de Berlín, Wilhelm von Waldeyer-Hartz (1836-1921), dio el nombre de «cromosomas» (también inventó la palabra «neurona» y un cierto número de términos embriológicos). Ahora bien, hay que señalar que Flemming fue incapaz de distinguir los cromosomas durante la interfase del ciclo celular, lo que dio lugar a interpretaciones diferentes del fenómeno. En cualquier caso, en 1885 Carl Rabl (1853-1917) señalaba que los gránuloscromosomas debían de mantenerse a lo largo de los diversos cambios que se producen durante la división celular, lo que le llevó en 1887 a defender su individualidad, un aspecto básico para la teoría cromosómica, aunque sería Theodor Boveri (1862-1915), de la Universidad de Múnich, quien clarificó y sistematizó esa teoría durante sus estudios citológicos: «En todas las células que proceden de la división de un huevo fecundado —escribía en 1891—, la mitad de los cromosomas son de origen paterno y la otra mitad, de origen materno».
Hugo de Vries.

Carl Correns.

Francis Galton, retrato de Frank Carter.
El propio Weismann llegó a situar su plasma germinal en el núcleo, aunque no estaba seguro de cuál era el papel que desempeñaban en los procesos hereditarios los filamentos nucleares —es decir, los cromosomas— observados. Por supuesto, la naturaleza de esa sustancia era desconocida, y continuaría siéndolo durante mucho tiempo (hasta la identificación del papel del ADN), pero la idea estaba clara, y así no nos debe extrañar que se introdujese un nuevo término-idea para describirla. En efecto, en 1909, en un libro titulado Elemente der Exakten Erblichkeitslehre (Elementos para el estudio de los caracteres hereditarios exactos), el danés Wilhelm Johannsen (1857-1927), catedrático de botánica en la Universidad de Copenhague, acuñaba el término gen (derivado de la raíz griega —que se encuentra también en vocablos como «génesis» o «generación»— que denota el concepto de engendrar), para denominar unas «partículas» hipotéticas —algo discreto— que portarían los cromosomas y que determinarían la herencia (todavía en 1926, y aunque él mismo había efectuado contribuciones sustanciales en favor de la identificación de los genes como entidades dotadas de realidad, Thomas Morgan, al que me referiré más adelante, recalcaba ese carácter hipotético de los genes en su libro The Theory of the Gene [Teoría del gen]: «Con el mismo criterio con el que los químicos dan por reales átomos invisibles y los físicos electrones, el estudioso de la herencia apela a unos elementos invisibles llamados genes»).

August Weismann.

Eric von Tschermak.

Diagrama de la mitosis celular según Walther Flemming (Zellsubstanz, Kern und Zelltheilung, 1882).

Wilhelm Johannsen y William Batenson ante la casa de este último (The Manor House, Merton).
Con el redescubrimiento del trabajo de Mendel y el apoyo de la teoría cromosómica, la genética pasó a ser uno de los campos más importantes de la investigación en biología. Estableciendo un paralelismo no demasiado correcto, aunque sugerente, se podría decir que la teoría cromosómica fue para la biología lo que el modelo atómico de Ernest Rutherford (1871-1937) para la física: aunque el átomo que el físico neozelandés instalado entonces en Manchester propuso en 1911 permitía explicar algunos fenómenos (como la difusión de partículas α), no aportaba nada a otros problemas —como el de los espectros asociados a los diferentes elementos—; para avanzar en esta dirección hubo que esperar un par de años: al modelo de Niels Bohr, que ya tomaba en consideración los cuantos de Planck-Einstein. Análogamente —aunque, insisto, este paralelismo tiene límites evidentes—, la teoría cromosómica permitía explicar algunos aspectos de la herencia, pero nada decía sobre cuestiones como los cambios que ahora asociamos a mutaciones, o a los mecanismos de la vida del tipo de, por ejemplo, la relación entre los genes («unidades estructurales», como veremos, más básicas localizadas dentro de los cromosomas) y las características propias del individuo.

Thomas Morgan trabajando en su laboratorio (en el «Cuarto de las moscas») de la Universidad de Columbia, c. 1915-1920.
En efecto, junto al problema de la composición de los cromosomas y de esas misteriosas unidades bautizadas como genes (y el de la posible relación existente entre ambos), estaba el de cómo es posible que se produjesen los cambios evolutivos tan radicales que terminaban conduciendo a la creación de nuevas especies, tal como defendía Darwin.
Las mutaciones constituyen, como sabemos, el motor responsable de tales cambios evolutivos, el mecanismo que contrarresta la estabilidad de la herencia. Pero la historia del concepto de mutación es larga, y confusa, ya que se empleó con significados diferentes. Hugo de Vries fue uno de los que primero recurrieron a él. En 1901, por ejemplo, la definía de la manera siguiente en el primer tomo de su opus magnum, Die mutationstheorie (La teoría de mutaciones; 1901-1903): «Por teoría de la mutación designo la aseveración de que las propiedades de los organismos están construidas por unidades acusadamente distintas… Los grados intermedios, que en tan gran número se dan entre las formas exteriores de plantas y animales, no existen entre estas unidades en mayor grado que entre las moléculas químicas». Sin embargo, y no obstante lo moderno que nos puedan sonar tales declaraciones, su teoría de la mutación no se amoldaba a la selección de las especies de Darwin. La teoría darwiniana, escribió, «supone que la variabilidad usual, la denominada individual, es el punto de partida del origen de especies nuevas. Según la teoría de la mutación, las dos variabilidades (la individual y la mutacional) son completamente independientes. Como espero demostrar, la variabilidad habitual no puede conducir a una transposición efectiva de los límites de la especie, incluso con la selección más intensa y rápida». Para él, «una especie nueva se origina de forma repentina. Surge de la ya existente sin preparación visible y sin transición». Desde semejante punto de vista, conceptos como selección natural o lucha por la existencia eran claramente superfluos para De Vries.
Serían, sobre todo, los estudios de Thomas Hunt Morgan (1866-1945), que se había doctorado en 1890 en la Universidad Johns Hopkins, los que representaron un momento crucial en la historia de la teoría de la mutación y, en general, de la genética.
Ahora bien, cuando Morgan llegó, en 1904, a la Universidad de Columbia, en Nueva York, como catedrático de Zoología experimental, era muy escéptico con respecto al mendelismo, esto es, a una teoría de la herencia que utilizase unidades discretas. Su cambio de actitud se debió básicamente a las investigaciones que emprendió con Drosophila melanogaster, la mosca del vinagre o de la fruta.
Inicialmente, Morgan había intentado estudiar la transmisión de caracteres en ratones y ratas, pero no tuvo éxito. Sólo hizo progresos tras elegir como protagonista de sus investigaciones a la humilde Drosophila, que podía crearse por millares en botellas de leche (como ventajas adicionales estaba el que el ciclo vital completo de estos animales, que únicamente poseen cuatro cromosomas, dura solamente diez días; además, los costes de manutención se reducían a unos pocos plátanos). Dadas estas características, pudo realizar sus investigaciones genéticas en una pequeña habitación de 4 × 6 metros, conocida como el «Cuarto de las moscas».
Entre 1907 y 1917, Morgan y su equipo trabajaron en el «Cuarto de las moscas» buscando mutaciones en ellas. Con un equipo ridículamente simple, el «Cuarto de las moscas» proporcionó poco a poco nuevos cimientos a la biología evolutiva. Los resultados que le llevaron a creer en las ideas mendelianas fueron los siguientes: aun cuando no se produjeron en sus cultivos notables mutaciones a nivel de especie, en 1910 encontró que en una de sus botellas de cría apareció repentinamente una curiosa mosca macho de ojos blancos. Morgan cruzó al macho mutante con una hembra normal (de ojos rojos). Todos los descendientes mostraron el rasgo normal, esto es, los ojos rojos. Sin embargo, cuando cruzó algunos miembros de la primera generación entre sí, descubrió que el carácter de los ojos blancos aparecía de nuevo, aunque sólo en los machos, nunca en las hembras. Por otra parte, si se cruzaba a un macho de ojos blancos con hembras de la primera generación, la mitad de los descendientes machos y la mitad de las descendientes hembras tenían ojos blancos. Morgan encontró que estos resultados se podían explicar sin demasiada dificultad a partir de la teoría mendeliano-cromosómica. Este fue el hecho que estimuló su transformación de escéptico mendeliano en partidario suyo.
No fue la menor de las aportaciones del grupo de Morgan el que fueran también capaces de encontrar marcadores cromosómicos que les sirvieron para establecer algo así como una cartografía de la estructura de los cromosomas: en 1911 construyeron el primer mapa cromosómico, que puso en evidencia la posición relativa de cinco genes ligados al sexo; once años después, otro de sus mapas indicaba las posiciones relativas de más de dos mil genes pertenecientes a los cromosomas de Drosophila. Con todo ello pudieron defender la tesis de que los genes se encuentran ordenados linealmente en los cromosomas y que experimentan cambios repentinos permanentes, o mutaciones, que producen un cambio en un rasgo concreto determinado por el gen, tal como el cambio del color rojo de un ojo al blanco.
Visto retrospectivamente, inmersos como estamos en una época en la que los genes son protagonistas indiscutidos de la biología contemporánea, el valor de los hallazgos de Morgan y su equipo adquiere dimensiones casi gigantescas. Una lectura superficial de su libro de 1926, todo un clásico de la ciencia contemporánea, The Theory of the Gene, en el que sintetizó las principales ideas a que había llegado tras décadas de investigaciones, muestra con claridad la importancia de su obra, y al hilo de ella, cómo había cambiado la genética desde los tiempos de Mendel, cuando apenas había transcurrido medio siglo de la publicación de «Versuche über Pflanzen-Hybriden», y hasta qué punto manejaba ideas que se instalarían definitivamente en el corpus de la biología. «Tenemos ahora —escribía— la posibilidad de formular la teoría de los genes.» Y como primer punto de esa teoría incluía el siguiente: «Los caracteres del individuo están relacionados a partes de elementos —genes— en la materia germinal, mantenidos juntos en un número de grupos enlazados».
Una cuestión de especial interés era la naturaleza de los genes. «¿Cómo puede ser que —se preguntaba—, tratándose de moléculas orgánicas, se conserven invariables, con plena estabilidad? Tal estabilidad significa que el gen es individualizado como una molécula viva, la cual se mantiene prácticamente invariable a pesar de su metabolismo, o que tiende a evolucionar según un modo definido… Hace unos años me propuse calcular el tamaño de los genes con la esperanza de que se consiguieran resultados útiles para avanzar en estos problemas. No contamos, sin embargo, con medidas suficientemente exactas para que podamos escapar de razonamientos que no sean puramente especulativos. Parece, sin embargo, que el volumen del gen es del orden de las mayores moléculas orgánicas. Podría tratarse, en efecto, de una gran molécula, pero es más verosímil pensar que el gen sea una estructura, un conjunto de moléculas orgánicas relacionadas por vínculos químicos, por la simple afinidad —como es una combinación química—, si no por otras fuerzas organizadoras.» Y añadía: «Es difícil, con todo, evitar la fascinante idea de que el gen sea constante precisamente porque constituya una entidad química organizada». Frases todas estas que suenan familiares, en absoluto extrañas, a todos aquellos que conocen lo que la biología molecular dice en la actualidad de los procesos hereditarios.
Fruto de sus trabajos, que tanto hicieron por la genética, en 1933 Morgan recibió el premio Nobel de Fisiología o Medicina, por «sus descubrimientos relativos a la función de los cromosomas en la transmisión de la herencia». Por entonces ya era, desde 1928, director del Laboratorio de Ciencias Biológicas del California Institute of Technology, en Pasadena, puesto que mantuvo hasta su muerte.
El desarrollo de la genética fue abriendo —con lentitud, a veces incluso entre contradicciones y puntos de vista opuestos— el camino a grandes avances en la comprensión de la vida, hasta tal punto que si del siglo XIX se puede decir que, en lo que se refiere a las ciencias biomédicas, fue el siglo de la fisiología, el XX fue el de la genética. En el plano teórico, suministró una base firme para entender la evolución. Al ser la fuente principal de novedad biológica, la mutación de genes se erigía en el motor que la gobernaba. De hecho, lo que selecciona el mecanismo darwiniano son organismos que transportan nuevos genes, o nuevas combinaciones de viejos genes, que confieren mayores ventajas en la lucha de la vida y la reproducción. Ahora bien, durante la primera mitad del siglo XX, mientras la genética se convertía en la reina de las ciencias biológicas, la naturaleza física del concepto central, el gen, continuó envuelto en misterio. Todavía en 1950, en un artículo titulado «The development of the gene theory» («El desarrollo de la teoría de los genes»), Hermann J. Muller (1890-1967), en el pasado colaborador de Morgan, y entonces uno de los más respetados genéticos y defensor de la teoría de los genes, declaraba que nadie sabía de qué estaban hechos los genes, cómo podían ser capaces de imponer su cáracter sobre los organismos que los transportan, o cómo se reproducen fielmente en la división celular.
No obstante, seis años antes de que Muller realizase semejantes afirmaciones, en 1944, Ostwald Avery (1877-1955) y sus colegas, Colin MacLeod (1909-1972) y Maclyn McCarty (1911-2005), habían anunciado en un artículo titulado «Induction of transformation by a desoxyribonucleic acid fraction isolated from Pneumococcus type III» («Inducción de transformación por una fracción de ácido desoxirribonucleico aislada del neumococo tipo II»), publicado en el Journal of Experimental Medicine, que los genes se encuentran sumergidos en ácido desoxirribonucleico (ADN). La razón de esta omisión por parte de Muller se debe a que, como en el caso de Mendel, el descubrimiento de Avery y sus colaboradores fue prematuro, en el sentido de que planteaba problemas importantes para cuya solución no se disponía aún de los recursos necesarios. Las ideas entonces en vigor acerca de la naturaleza molecular del ADN hacían que fuese muy complicado pensar que el ADN pudiese ser el vehículo de la información hereditaria. Lo que desde luego no fue similar al caso de Mendel es que el artículo de Avery, MacLeod y McCarty tuviese que ser redescubierto, ya que su publicación constituyó toda una sensación, y sin duda Muller lo conocía muy bien cuando escribió su artículo.

Aparato de rayos X para inducir mutaciones, siendo utilizado por Clarence Oliver en la Universidad de Texas (1927).
Uno de los motivos por los que se puede calificar —como he hecho— el descubrimiento de Avery de «prematuro» es porque no basta con encontrar «algo» —ADN, en este caso— en un lugar (los cromosomas) para establecer una relación causal explicativa. Para hacerlo es preciso elaborar algún tipo de mecanismo, «abrir», por así decirlo, «la caja negra». Y en aquel momento y para aquel problema, la biología no era suficiente, se necesitaba el auxilio de otras disciplinas científicas, más dotadas para la construcción de «mecanismos». Disciplinas como la química (la bioquímica fue —y es— imprescindible en el desarrollo de la biología molecular) y la física.
Antes de continuar por esta línea, es oportuno aprovechar la ocasión para señalar que aunque tendemos a establecer fronteras definidas entre las diferentes ciencias (esto pertenece al dominio de la química, esto al de la matemática, aquello al de la biología, eso al de la física, etc.), con frecuencia se necesitan herramientas de diversas disciplinas para resolver problemas vinculados a una de ellas. Es la interdisciplinariedad, con la que ya nos hemos encontrado, explícita o implícitamente (recordemos el caso de la fisiología del siglo XIX), en otros lugares de este libro. De hecho, se puede argumentar que una de las consecuencias del desarrollo científico es el derrumbamiento de tales fronteras, surgiendo disciplinas mixtas —como químico-física, bioquímica, biofísica o electroquímica, sin olvidar las «tecnocientíficas»—; más aún, algunas de las novedades científicas más atractivas que han surgido a lo largo del último medio siglo tienen su razón de ser precisamente en la interdisciplinariedad. El caos, esa sensibilidad extrema a las condiciones iniciales que hace que sea imposible predecir la evolución futura de un sistema, constituye un buen ejemplo. Su descubridor fue Edward Lorenz (1917-2008), un meteorólogo estadounidense con una sólida formación matemática. El artículo de 1963 en el que Lorenz introdujo el caos trataba de un fenómeno típicamente atmosférico: las corrientes de convección. Sin embargo, investigaciones posteriores han mostrado que existen movimientos caóticos en muchos otros dominios: en la física, por ejemplo, en la magnetización del helio superfluido o en los plasmas; en la ingeniería, en numerosos fenómenos aerodinámicos; en medicina, en oscilaciones cardíacas, como las arritmias, también en la dinámica del funcionamiento cerebral; en ecología, en ciertos modelos que estudian las relaciones entre depredador y presa. Tampoco se libra del caos (en un sentido matemático) la economía, en donde está siendo estudiado su posible efecto en macromodelos.

Ostwald Avery en su laboratorio del Rockefeller Hospital.
Pero regresemos a la biología y al problema de los mecanismos de transmisión de los caracteres hereditarios. Como apuntaba antes, este problema tuvo que esperar para su resolución, como veremos a continuación, a una línea de investigación muy influida por la física —por no decir estrechamente ligada a ella—. En la actualidad, que fuese así —que cromosomas, genes o ADN se explicasen en términos físicos o químicos— nos puede parecer trivial, no absolutamente necesario pero sí plausible. Sin embargo, en las primeras décadas de «existencia» de los genes, todavía era común aceptar de manera acrítica fronteras entre disciplinas, permaneciendo la genética separada habitualmente de las ciencias físico-químicas. Así, en 1939 en un libro de texto, Introduction to Genetics (Introducción a la genética), dos biólogos del California Institute of Technology, discípulos de Morgan, Alfred H. Sturtevant (1891-1970) y George W. Beadle (1903-1989), escribían: «La física, la química, la astronomía y la fisiología tratan con átomos, moléculas, electrones, centímetros, segundos, gramos (todos sus sistemas de medidas se reducen a estas unidades comunes). La genética no tiene a ninguno de estos como componentes de sus unidades fundamentales y, sin embargo, es una materia formulada matemáticamente que es lógicamente completa y autocontenida».
Física, biología y mutaciones
Para introducir la línea de investigación que terminaría relacionando la física con la genética, me voy a referir de entrada a un científico alemán, que iba para físico cuántico: Max Delbrück (1906-1981).
Tras haber estudiado física, Delbrück se planteó doctorarse, recibiendo un tema de Walter Heitler (1904-1981). Teniendo en cuenta de quién procedía la idea (en 1927, Heitler fue el autor, en colaboración con Fritz London [1900-1954], de un influyente artículo que marcó el desarrollo de la teoría del enlace químico, titulado «Interaction between neutral atoms and homopolar binding according to quantum mechanics» [«Interacción entre átomos neutros y enlace homopolar según la mecánica cuántica»]), no sorprende que el tema fuera explicar en términos semicuantitativos por qué el enlace covalente entre dos átomos de litio era mucho más débil que el enlace homólogo formado entre dos átomos de hidrógeno. En 1929, en Gotinga, Delbrück finalizó su tesis, que involucraba cálculos bastante engorrosos.
Tras pasar un año en Bristol, con John Lennard-Jones (1894-1954), Delbrück obtuvo una beca de la Fundación Rockefeller por un año. Decidió pasar la primera mitad en Copenhague, junto a Niels Bohr, y la segunda en Zúrich (llegó allí en febrero de 1931), con Wolfgang Pauli (1900-1958), uno de los «grandes» de la historia de la mecánica cuántica. Como muchos otros científicos de la época, Delbrück pasó a formar parte del círculo de Bohr, y precisamente debido a este hecho recibió una influencia decisiva: durante una conferencia que Bohr pronunció el 15 de agosto de 1932, titulada «Luz y vida». En uno de sus artículos, que escribió muchos años después, Delbrück se refirió a aquella conferencia de la manera siguiente:
Con respecto a la biología, en su conferencia «Luz y vida» de 1932, Bohr propuso que tal vez no fuera reducible a la física atómica. Sugirió que entre los aspectos físicos y psicológicos de la vida acaso hubiera una complementariedad, análoga a los aspectos de onda y de partícula del electrón. En ese caso habría una especie de principio de incertidumbre de la biología…
Desde el punto de vista de la física, en aquella época los misterios de la vida eran, en efecto, impresionantes. Los fisiólogos celulares habían descubierto innumerables modos con los que las células responden inteligentemente a cambios de las condiciones ambientales. Los embriólogos habían demostrado cosas tan asombrosas como la posibilidad de obtener dos animales a partir de dos mitades de un embrión. Estos descubrimientos recordaban vagamente la «unidad orgánica» del átomo y la estabilidad de sus estados cuánticos. La estabilidad de los genes y el álgebra de la genética mendeliana sugirieron a Bohr que los procesos que subyacían a los fenómenos eran parecidos a la teoría cuántica.
Vemos así que la idea de que la física cuántica tenía algo que decir con relación al desarrollo de la biología data, al menos, de comienzos de la década de 1930. El propio Delbrück se dedicó, a partir de 1934, a buscar una conexión entre la biología y la física, y lo hizo de una manera muy directa, interesándose por la influencia de la luz en la vida. Una ayuda esencial en semejante dirección surgió de la genética clásica: en 1927, Hermann Muller, con quien ya nos hemos encontrado, describió una técnica por la que los rayos X inducen mutaciones en la mosca Drosophila en una proporción mucho mayor que la que se produce de forma natural. Además de permitir a Muller aislar varios cientos de mutaciones, una gran parte de las cuales se mantuvieron en las siguientes generaciones de moscas, este descubrimiento abría el camino a los físicos para que estudiasen los procesos físico-químicos en la entidad biológica denominada «gen», concepto que el propio Muller promovió con fervor, postulando que era la auténtica «base de la vida», aunque se daba cuenta de que «el genetista es incapaz de analizar… sus propiedades» (1936).
Muller urgió a los físicos para que fuesen en su ayuda, y de hecho en 1932 se trasladó a Berlín para colaborar con un emigrado ruso, Nikolai Timoféeff-Ressovsky (1900-1981), que dirigía el Laboratorio de Genética del Instituto de Investigación del Cerebro de la Asociación Káiser Guilllermo. Por entonces, Delbrück se encontraba también en Berlín, en donde había organizado un pequeño grupo de discusión en la casa de su madre. Allí conoció a Timoféeff-Ressovsky. Con él y con Karl Günter Zimmer (1911-1988), desarrolló en 1935 un modelo cuántico de mutación genética («Über die Natur der Genmutation und der Genstruktur» [«Sobre la naturaleza de la mutación genética y la estructura de los genes»]), que aunque resultó incorrecto, a la postre ejerció una notable influencia. De hecho, se puede decir que aquel trabajo constituye una de las primeras aproximaciones a una auténtica biología molecular.

Max Delbrück en el California Institute of Technology.
Si el artículo de Delbrück, Timoféeff-Ressovsky y Zimmer ejerció alguna influencia, fue debido a la labor de divulgación ejercida en favor suyo por un librito del creador de la mecánica cuántica ondulatoria, el físico y premio Nobel austríaco Erwin Schrödinger (1887-1961): What is Life? (¿Qué es la vida?), publicado en 1944 y fruto de una serie de conferencias en el Institute for Advanced Studies de Dublín, al que Schödinger se había incorporado tras exiliarse de la Alemania hitleriana.
Que el texto de Schrödinger participaba de un espíritu análogo al de Delbrück es algo que se puede comprobar fácilmente sin más que ojear sus páginas, en las que se pueden encontrar sentencias como la siguiente: «El holandés Hugo de Vries descubrió hace unos cuarenta años que incluso en la descendencia de cepas realmente puras un número muy pequeño de individuos, algo así como dos o tres entre varias docenas de miles, aparece con cambios pequeños, pero que suponen una especie de “salto”. La expresión “salto” no quiere significar que el cambio sea especialmente importante, sino que supone una discontinuidad, en el sentido de que no hay formas intermedias entre la forma inalterada y los pocos individuos que han cambiado. Tras su observación, De Vries les dio el nombre de mutaciones. El hecho significativo es la discontinuidad. Al físico le recuerda la teoría cuántica, según la cual no hay energías intermedias entre dos niveles energéticos contiguos. Podríamos denominar a la teoría de la mutación, de forma figurada, la teoría cuántica de la biología… Las mutaciones se deben, de hecho, a saltos cuánticos en las moléculas del gen. Pero la teoría cuántica sólo tenía dos años cuando De Vries publicó su teoría de la mutación, en el año 1902. No es, pues, extraño que se necesitase una generación más para descubrir la íntima relación entre ambas».

Erwin Schrödinger.
Dan una idea del impacto de What is Life? las siguientes palabras de Francis Crick, quien, por cierto, se había graduado en física y trabajado como físico antes de iniciarse en la biología, y al que el libro de Schrödinger influyó considerablemente: «Leí el librito… de Erwin Schrödinger What is Life?», escribió Crick en un libro autobiográfico (What Mad Pursuit [Que loco propósito]; 1989), refiriéndose a los años de la Segunda Guerra Mundial (que él pasó trabajando para el Ministerio de Defensa británico), aunque «sólo más adelante logré ver sus limitaciones —como la mayoría de los físicos, su autor no sabía nada de química—, pero sin duda creaba la sensación de que cosas importantes estaban a la vuelta de la esquina».
En concreto, las principales preguntas que se hizo Schrödinger en What is Life? se podrían resumir de la siguiente manera: ¿Cuál es la estructura física de las moléculas que se duplican cuando se dividen los cromosomas? ¿Cómo se debe entender el proceso de la duplicación? ¿Cómo retienen, de generación a generación, estas moléculas su individualidad (problema de la herencia)? ¿Cómo logran controlar el metabolismo de las células? ¿Cómo crean la organización que se ve en la estructura y en las funciones de los organismos superiores? A Schrödinger le preocupaba, en suma, el hecho de que los acontecimientos a nivel atómico son muy inestables y efímeros, mientras que los organismos vivos muestran una gran estabilidad. No contestó las preguntas que se formuló, limitándose a sugerir que, de alguna manera, la información necesaria para retener los caracteres que se transmiten de generación en generación debía de estar contenida en los enlaces covalentes de un «cristal aperiódico gigante», que denominó «el código hereditario», pero el simple hecho de formular semejantes cuestiones contribuyó a favorecer el desarrollo de la biología molecular.
Mientras que Schrödinger poco más podía añadir al problema de la vida, o de los genes, Delbrück ya había comenzado antes (al menos en 1938) a estudiarlos experimentalmente, instalado en Estados Unidos, en el California Institute of Technology, recurriendo a los virus bacterianos, o fagos.
Aunque los fagos son demasiado pequeños para observarlos con microscopios ordinarios, y estructural y químicamente son muy sencillos (mitad proteínas, mitad ADN), están dotados de una gran capacidad de autorreproducción. Delbrück demostró que un fago que infecta a una célula puede dar lugar a cientos de fagos idénticos en media hora, con lo que se convertían en extraordinarios instrumentos para estudiar la replicación genética. Dos años después, se encontró con Salvador Luria (1912-1991) y Alfred Hershey (1908-1997), formando el conocido «Grupo de los Fagos», cuyos miembros estaban unidos por un deseo común: resolver el misterio del gen. Es interesante también mencionar que en 1947, Luria, entonces profesor en Indiana, tomó como estudiante graduado a un joven llamado James Watson, iniciándole como miembro del Grupo de los Fagos. La «conexión física del ADN», esta vez, ciertamente, de manera algo más indirecta que en el caso de Crick, se cerraba.

Martha Chase y Alfred Hershey en Cold Spring Harbor.
Pero antes de abandonar esta senda, la de los fagos, mencionaré la importancia que tuvieron para la aceptación del ADN como material genético los experimentos realizados en 1952 por otros miembros de ese grupo, el citado Alfred Hershey y Martha Chase (1927-2003), con los que demostraron que cuando un fago (el denominado T2) infecta a una bacteria y se reproduce allí produciendo más fagos, solamente el ácido nucleico, el ADN, entra en la bacteria, mientras que la proteína del fago permanece en el exterior. Estos resultados resaltaron la importancia del ADN, reavivando asimismo el interés y el recuerdo de los de Avery de 1944, facilitando las nuevas observaciones que Watson y Crick realizarían enseguida. En su autobiografía, Salvador Luria escribió, en este sentido:
En un conjunto de experimentos justamente célebres por su elegante sencillez, [Alfred Hershey] y su colega Martha Chase demostraron que cuando una partícula de bacteriófago, que se compone de una proteína y ADN, ataca una bacteria, la proteína queda fuera y el ADN penetra en la bacteria, se multiplica y hace surgir más bacteriófagos. En los bacteriófagos, como en las bacterias de Avery y como en cualquier organismo vivo, hoy estamos seguros de que los genes son ADN (algunos virus se las arreglan con genes hechos de otra clase de ácido nucleico llamado ARN).
La importancia de los descubrimientos que identificaron como ADN el material genético, primero de las bacterias, luego de los bacteriófagos y, a la postre, de todos los organismos consistió en que convirtieron el problema del gen en un problema para la química (aunque seguía siendo, plenamente, un problema de genética). Aún no se conocía la estructura del ADN a nivel molecular, pero al menos químicos y genéticos sabían que estaban tratando con una sustancia química específica.
La importancia de las técnicas de difracción de rayos X
Las ideas procedentes de la física relativas al concepto de mutación y de cómo provocarlas fueron importantes, pero en modo alguno se puede explicar sólo a partir de ellas el universo de los conocimientos biológicos que poseemos en la actualidad, en los que la estructura del ADN ocupa un lugar destacado. Es inevitable referirse también a las técnicas de difracción de rayos X, un instrumento, surgido asimismo en el seno de la física, esencial para la determinación de estructuras moleculares.
Los trabajos de investigación y, sobre todo de promoción, de William Lawrence Bragg (1890-1971), mientras fue director del Laboratorio Cavendish de Cambridge (entre 1938 y 1954), resultaron especialmente importantes en este sentido (William Lawrence y su padre, William Henry Bragg [1862-1942], habían desarrollado a partir de 1913 las técnicas básicas de difracción de rayos X para analizar la estructura molecular de cristales de todo tipo de sustancias). Bajo la dirección de W. L. Bragg el inhóspito laboratorio de cristalografía del Cavendish fue creciendo, hasta convertirse en uno de los centros más importantes del mundo.

John Kendrew (izquierda) y Max Perutz (derecha) ante un modelo de hemoglobina de 1962, enmarcados en un ordenador.
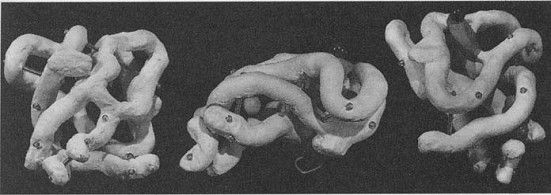
Modelo de hemoglobina desarrollado por John Kendrew.
Uno de los investigadores que estaban en el Cavendish cuando llegó Bragg era Max Perutz (1914-2004), un joven químico austríaco que se había trasladado a Cambridge. En Viena, Perutz había podido conseguir unos cristales de hemoglobina de caballo (la proteína que da color rojo a las células de la sangre); pronto se dio cuenta de que tenían «la estructura cristalina más sencilla de todas las proteínas de peso molecular comparable, con características que hacen saltar de gozo al corazón del cristalógrafo». Sin embargo, una cosa era que fuera más sencilla que otras macromoléculas y otra que fuese asequible resolver el problema de su estructura; ante la ausencia de un método directo para obtener las posiciones atómicas, parecía imposible intentar abordar una estructura molecular tan compleja como la de la hemoglobina. Era —en palabras del propio Perutz— «tan prometedor como un viaje a la Luna». Sin embargo, continuó con aquel proyecto, que terminaría dando fruto en 1953, cuando Perutz obtuvo imágenes que demostraban que, en principio, la estructura de la molécula de hemoglobina podía, efectivamente, ser desentrañada. Se tardaron otros cuatro años más en dar con el modelo definitivo, siendo de nuevo Perutz el responsable, quien en 1962 —el mismo año en que fueron galardonados, en Medicina, Crick, Watson y Wilkins— recibió el premio Nobel de Química por sus «investigaciones sobre la estructura de proteínas globulares (en particular la hemoglobina)». Lo compartió con otro investigador del Cavendish, John C. Kendrew (1917-1997), a quien el propio Perutz había enseñado los fundamentos de la cristalografía, que lo recibió por «la determinación de la estructura de la mioglobina».

John D. Bernal fotografiado por Lettice Ramsey en Cambridge (1932).
Habida cuenta de estos hechos, no debe sorprendernos que fuese también en el Cavendish donde se descubrió, en 1953, la estructura en doble hélice del ácido desoxirribonucleico, a la que me referiré enseguida.
Sobre el reduccionismo, Biología versus física y química:
Los comentarios precedentes, relativos al papel que la física ha desempeñado en el desarrollo de la biología molecular (comentarios que se pueden hacer extensivos, en otros apartados, a la química, como veremos a continuación), pueden inducir a algunos a pensar que la vieja ecuación reduccionista, «la biología se reduce a la química, la química a la física, luego la biología (esto es, la vida) se reduce a la física», es cierta. Semejante ecuación-planteamiento es, no obstante, tan limitado y simplista como burdo y muestra una idea muy primitiva de lo que es la ciencia. Es evidente que el conocimiento de, por ejemplo, las estructuras atómicas que proporciona la física es instrumento esencial para comprender el enlace químico, y que éste es, a su vez, imprescindible para entender el funcionamiento de la materia biológica, pero la biología —al igual que cualquier otra ciencia— es más que eso. Posee problemas y técnicas propias, y es heredera de una tradición y de unos estilos que se han ido configurando a lo largo de siglos y sin los cuales no sería lo que es. Ni la investigación, ni las cuestiones que se reconocen como legítimas, ni tampoco las instituciones y procedimientos que la configuran como disciplina científica permanecen —pueden permanecer— al margen de semejantes factores.
Aunque no recoge todos estos aspectos, la siguiente cita trata de este asunto. Su autor es el célebre cristalógrafo británico y militante del socialismo científico (fue marxista), John Desmond Bernal (1901-1971). La formación de Bernal, sus intereses científicos (realizó, desde la cristalografía y difracción de rayos X, notables aportaciones a la biología molecular, disciplina de la que, de hecho, fue uno de sus «padres fundadores»), históricos y sociales (entre sus obras figura el famoso libro, The Social Function of Science [1939]) dan especial relieve a sus opiniones.
La biología ha recibido nuevas contribuciones de la química y de la física. La nueva comprensión del comportamiento de las unidades más pequeñas de la materia —los átomos y las moléculas—, lo mismo que las técnicas para estudiarlas, han sido de un valor inestimable para la biología. Pero esto no significa, como algunos están dispuestos a pensar, que la biología se haya convertido en una rama de la física y la química. Por el contrario, el empleo de los conocimientos físicos o químicos para explicar los aspectos mecánicos, eléctricos o químicos de los organismos vivos, ha servido sólo para destacar más todavía sus aspectos biológicos. A pesar de que se pueden describir en términos físicos, estos fenómenos no tienen lugar mediante mecanismos hechos por algún artesano divino y conforme a modelos ideales establecidos para toda la eternidad, sino que son procesos que se regulan y se reproducen por sí mismos y cuya forma actual es el resultado de una evolución que ha durado millones de años.
John D. Bernal, Science in History, 1954 (La ciencia en la historia)
La conexión química: de la bioquímica a la biología molecular
Aunque he estado haciendo hincapié en la «conexión física» de las investigaciones que finalmente conducirían al descubrimiento de la estructura del ADN, también he aludido a que la genética —más estrictamente, la biología en su conjunto— es asimismo deudora de otras disciplinas científicas. Como la química.
De hecho, en un sentido estricto, si mi intención no hubiese sido la de centrarme en los mecanismos hereditarios, en la genética, debería haber empezado por señalar que no es posible comprender las transformaciones que ha experimentado la biología del siglo XX sin tomar en cuenta la colonización que ésta sufrió durante su primer tercio por la química, la química orgánica en especial, que se plasmó en la aparición de una nueva disciplina, la bioquímica, desarrollada sobre todo en Gran Bretaña y Alemania, y que se centró preferentemente en el estudio de moléculas pequeñas o intermedias. En el capítulo 5 me referí, con relación a la «medicina científica», a las aportaciones que la química realizó a la fisiología, pero aquello fue más una «reunión» que una verdadera «unión» a largo plazo (a pesar de que apareciesen incluso revistas —la primera en 1877— con el nombre de «Química fisiológica» en su encabezamiento), en el sentido de que no condujo al establecimiento de una nueva disciplina, o subdisciplina, auténticamente institucionalizada, como sería más tarde el caso con la bioquímica.
Aunque en modo alguno quiero decir que no aportase bastante a la genética, en don de la bioquímica desempeñó un papel absolutamente central fue en el estudio de los procesos metabólicos, en la purificación, aislamiento y caracterización de enzimas, cinética de las reacciones que éstas catalizan, y en su integración en la explicación de efectos fisiológicos, como la contracción muscular o el crecimiento de tejidos. En otras palabras, en el establecimiento, a mediados de la década de los años treinta, de lo que se ha denominado «metabolismo intermediario». (También se podría hablar de lo que la bioquímica aportó a Gran Bretaña y Alemania durante la Primera Guerra Mundial, desarrollando, por ejemplo, procesos para la producción de acetona, un ingrediente básico para la fabricación de municiones, que hasta entonces se obtenían de nitratos procedentes de minas chilenas, cuyo acceso quedó interrumpido por los bloqueos navales.)
Pero en esa misma década a la que me refería, la de 1930, las técnicas físicas procedentes sobre todo de la física atómica (como la difracción de rayos X, mencionada con anterioridad), prácticamente desconocidas por los bioquímicos y sin las cuales no era posible desvelar la estructura espacial de macromoléculas, como son las proteínas (las enzimas también son, recordemos, proteínas), dieron paso a una nueva fase de la biología contemporánea, la de la biología molecular (momentos importantes en este sentido fueron la primera fotografía de rayos X de una proteína —1934— y la primera teoría sobre la estructura de una proteína, en 1936). Este fue el camino que condujo al descubrimiento de la doble hélice de ADN.
La estructura del ADN
James Watson (n. 1928) y Francis Crick (1916-2004) fueron los autores de tal hallazgo, pero también hay que recordar a otros. Como Rosalind Franklin (1920-1958), quien demostró la existencia de dos formas de ADN, las denominadas A y B, y con gran esfuerzo determinó la densidad, las dimensiones celulares exactas y la simetría de la forma A, evidencia que sugirió muy firmemente que la estructura tenía dos cadenas (y no sólo una, como había pensado otro científico que habría que recordar, el químico cuántico estadounidense Linus Pauling [1901-1994], cuya hélice α inspiró a Watson y a Crick), que circulaban en direcciones opuestas. También sería necesario hablar de Maurice Wilkins (1916-2004), que compartió con Crick y Watson el premio Nobel de 1962, y que fue quien demostró que las imágenes de difracción de rayos X de fibras de ADN se ajustaban a variantes del modelo de doble hélice. En el lado químico, no habría sido posible construir modelos correctos sin la fórmula química general del ADN, determinada sobre todo por el trabajo de Alexander R. Todd (1907-1997), aunque la información más vital fue suministrada por las investigaciones de Erwin Chargaff (1905-2002), que encontró que en el ADN existía la misma cantidad de adenina que de tinina y la misma de guanina que de citosina, sustancias químicas fundamentales a las que volveré enseguida. Y, por supuesto, igualmente fundamental fue el artículo de Avery, MacLeod y McCarty donde demostraban que el factor de multiplicación del neumococo estaba hecho casi con seguridad de ADN. Otra clave importante fue suministrada por John Masson Gulland (1898-1947), cuyo trabajo en la curva de análisis volumétrico del ADN sugirió, aunque no demostró, que el ADN en solución se mantenía unido por enlaces de hidrógeno.

Maurice Wilkins.

Erwin Chargaff.

James Watson y Francis Crick fotografiados el 21 de mayo de 1953 por Antony Barrington Brown en el Laboratorio Cavendish ante un modelo tridimensional de ADN.
La importancia que algunos de estos trabajos tuvieron para el descubrimiento de Watson y Crick —una importancia que ha dado origen a más de una polémica sobre prioridades y méritos (especialmente en lo que se refiere a los trabajos de Rosalind Franklin)— se hacía ya evidente en su propia puesta en escena. Watson y Crick publicaron su hallazgo en un esplendorosamente breve (prácticamente sólo una página) artículo («Molecular structure of nucleic acids: A structure for deoxyribonucleic acid» [«Estructura molecular de los ácidos nucleicos: Una estructura para el ácido desoxirribonucleico»]), que apareció el 25 de abril de 1953 en la revista inglesa Nature (volumen 171, pp. 737-738). A continuación (pp. 738-740) aparecía otro («Molecular structure of deoxypentose nucleic acids» [«Estructura molecular de ácidos desoxipentosanucleicos»]) firmado por Maurice Wilkins y dos de sus colaboradores en el King’s College: Alexander R. Stokes (1919-2003) y Herbert R. Wilson (n. 1929). Finalmente (pp. 740-741), venía un tercero («Molecular configuration in sodium thymonucleate» [«Configuración molecular en el timonucleato de sodio»]) de Rosalind Franklin y Raymond G. Gosling (n. 1926).
En «Molecular structure of nucleic acids», Watson y Crick señalaban que el ADN está constituido por dos cadenas, formando una doble hélice, cada una formada por cuatro compuestos químicos (bases) —combinaciones de carbono, nitrógeno, oxígeno, hidrógeno y fósforo—, denominados adenina (A), guanina (G), citosina (C) y timina (T). La estructura de la cadena es muy sencilla: lo que se puede considerar como su esqueleto está formado por una alternancia regular de ácido fosfórico (P) y un azúcar, la desoxirribosa (D). Sobre este esqueleto, se superponen las cuatro bases mencionadas antes, pero de forma que a cada azúcar D va unida una base, A, C, G o T, formando una secuencia determinada. El ADN surge cuando se unen dos cadenas de este tipo, mediante el procedimiento de establecer uniones entre parejas de bases, pero sólo de manera que la T de una cadena se asocie a la A de la otra, o la G a la C. Cualquier otro emparejamiento está prohibido. Con semejante estructura se puede comprender fácilmente (en principio; hay que elaborar los detalles) el proceso de la duplicación del ADN en cada célula: en un momento dado de la vida de ésta, la doble hélice se escinde en dos segmentos helicoidales que, al captar en el medio en el que se encuentran el azúcar, el fosfato y la base, reconstruyen sus complementarios, de manera, además, perfectamente fiel, de generación a generación (salvo mutaciones), debido a la ley del emparejamiento de las bases. «No ha escapado a nuestra atención —escribían Watson y Crick en su artículo— que el emparejamiento específico que hemos postulado sugiere un posible mecanismo de copiado para el material genético.» Esto es todo lo que decían, en un prodigio de brevedad y concisión. ¡Pero era tanto lo que englobaban esas parcas frases!
La era de la biología molecular llegó entonces a su momento crucial, más característico e importante. Quedó claro finalmente que el ADN aparece en forma de filamentos muy largos en los cromosomas. Tan largos que si pudiésemos extender el ADN de una célula humana, formaríamos un hilo de unos tres metros de longitud. El «genoma» (conjunto de instrucciones —o de genes— que permiten construir un organismo) humano se encuentra en esos hilos, y consta de unos 3.000 millones de pares de bases. Entre las inmensas posibilidades de comprensión y actuación que abría el nuevo esquema, una era que desaparecía ya de manera definitiva, nítida, la posibilidad de que alguien dijese lo que había escrito, como vimos, Morgan: «El estudioso de la herencia apela a unos elementos invisibles llamados genes». Ahora un gen se podía definir como una secuencia de pares de bases a lo largo de un trozo de ADN, que tiene una función específica conocida.

Primera página del artículo sobre la estructura del ADN de Watson y Crick (Nature, 25 de abril de 1953).
El artículo fundacional de Watson y Crick en Nature fue datado por sus autores el 2 de abril y apareció publicado en el número del 25 de abril. Se trataba, como ya he indicado, de un escrito —una nota— extremadamente sucinto. Enseguida prepararon un artículo más extenso, en el que elaboraban sus explicaciones y planteamientos. A continuación reproduzco algunos pasajes de este trabajo, que apareció en el número de Nature del 30 de mayo.
J. Watson y F. Crick, «Genetic implications of the structure of deoxyribonucleic acid» («Implicaciones genéticas de la estructura del ácido desoxirribonucleico»), Nature 171, pp. 964-967 (1953):
La importancia del ácido desoxirribonucleico en las células vivas es indiscutible. Se encuentra en todas las células que se dividen, sobre todo, si no totalmente, en el núcleo, donde es un constituyente esencial de los cromosomas. Hay muchos indicios de que es el portador de una parte de (si no de toda) la especificidad genética de los cromosomas y por tanto del propio gen. Hasta ahora, no obstante, no se ha demostrado la manera como lleva a cabo la operación esencial que se exige al material genético, es decir, la de su duplicación exacta.
Nosotros hemos propuesto recientemente una estructura para la sal del ácido desoxirribonucleico que, si es correcta, sugiere inmediatamente un mecanismo para su duplicación. Los resultados obtenidos mediante rayos X por los investigadores del King’s College de Londres y presentados al mismo tiempo, apoyan cualitativamente nuestra estructura y son incompatibles con todas las estructuras presentadas anteriormente. Aunque nuestra estructura no puede considerarse totalmente verificada hasta que se haya realizado una comparación más amplia con los datos obtenidos por rayos X, tenemos ahora la suficiente confianza en su exactitud general para poder discutir sus implicaciones genéticas. Al hacer esto damos por sentado que las fibras de la sal del ácido desoxirribonucleico no son artefactos fruto del método de preparación, ya que ha sido demostrado por Wilkins y sus colaboradores que se obtienen modelos de rayos X similares tanto a partir de las fibras aisladas como a partir de ciertos materiales biológicos intactos, tales como cabezas de espermatozoos y bacteriófagos…
La primera característica de nuestra estructura que tiene interés biológico es que está formada por dos cadenas, no por una. Estas dos cadenas están enrolladas alrededor de un eje común…
La otra característica importante desde el punto de vista biológico es la forma en que las dos cadenas se mantienen unidas. Se realiza por medio de los enlaces de hidrógeno entre las bases… Las bases se unen a pares, una base de una cadena establece un enlace de hidrógeno con la otra cadena. El punto importante reside en el hecho de que sólo unos ciertos pares de bases se ajustan a esta estructura. Un miembro de cada par debe ser una purina y el otro una piramidina para poder establecer contacto entre las dos cadenas. Si un par estuviera formado por dos purinas, por ejemplo, no cabría en la estructura.
Nosotros creemos que las bases están presentes casi totalmente en su forma tautomérica más probable. Si esto es cierto, las condiciones para que se formen enlaces de hidrógeno son más restrictivas y los únicos pares de bases posibles son:
adenina con timina;
guanina con citosina
…
El esqueleto fosfato-azúcar de nuestro modelo es completamente regular, pero cualquier secuencia de pares de bases puede ajustarse a esta estructura. De aquí se deduce que en una molécula larga son posibles muchas permutaciones diferentes y, por lo tanto, parece probable que la secuencia particular de las bases sea el código que transporta la información genética. Si el orden real de las bases en una de las cadenas está determinado, se puede escribir el orden exacto de las bases de la otra cadena debido a su apareamiento específico. Así, una cadena es, como si dijéramos, el complemento de la otra y es esta característica la que sugiere la forma en que la molécula de ácido desoxirribonucleico puede duplicarse…
De momento el esquema general para la reproducción del ácido desoxirribonucleico debe considerarse como especulativo. Incluso si fuera correcto, es evidente por lo que hemos dicho que queda mucho por descubrir para que el proceso de la duplicación genética pueda describirse con detalle… ¿Qué es lo que hace que el par de cadenas se desenrollen y separen? ¿Cuál es el papel exacto de la proteína? ¿Está el cromosoma formado por un largo par de cadenas de ácido desoxirribonucleico, o lo está por porciones del ácido unidas entre sí por proteínas?
A pesar de todas estas incertidumbres creemos que la estructura que hemos propuesto para el ácido desoxirribonucleico puede ayudar a solucionar uno de los problemas biológicos más fundamentales: la base molecular del molde necesario para la replicación genética. La hipótesis que sugerimos es que el molde es el modelo de bases formado por un cadena de ácido desoxirribonucleico y que el gen contiene un par complementario de estos moldes.
Una nueva era sociocientífica
Visto retrospectivamente, se puede pensar que el descubrimiento de la estructura del ADN abrió inmediatamente una nueva era, que podría denominarse de la biología molecular, en la que todavía nos encontraríamos. No está, sin embargo, tan claro que semejante caracterización sea la más adecuada. No hay duda que el hallazgo de Watson y Crick creó instrumentos conceptuales sin los cuales no habrían sido posibles los cambios —reales o esperados— que configuran aspectos centrales del mundo en que vivimos, pero también hay que tener en cuenta que para que comenzaran a aparecer en el horizonte esas transformaciones fue necesario más que las herramientas habilitadas por los resultados anunciados en 1953. Y es que no siempre —más aún, acaso raras veces (pero esta es otra cuestión que habría que estudiar en sí misma)— los resultados científicos, aunque sean revolucionarios, poseen la suficiente capacidad de transformación como para afectar drásticamente a la sociedad.
En el caso del que me estoy ocupando ahora esto quiere decir lo siguiente: por un lado se tiene que la determinación de la estructura molecular del ADN permitió comprender, como nunca antes se había logrado, los mecanismos que rigen los procesos hereditarios, sobre cuya importancia es innecesario detenerse; por otro, sabemos —lo leemos, escuchamos o vemos constantemente en los medios de comunicación— que nos encontramos en medio, o en los albores, de una gran revolución tecnocientífica que, se dice, cambiará nuestras vidas (o las expectativas que todo humano se forma de su futuro vital), con la enorme implicación socioeconómica —y a la postre política— que ello implica. No es extraño que de las dos manifestaciones anteriores la mayoría de las personas concluya que la ecuación básica es algo así como: una vez disponibles los resultados de Watson y Crick de 1953, lo fundamental estaba hecho, siendo cuestión de tiempo que se explotasen sus consecuencias.
En la medida en que la identificación de la doble hélice es condición necesaria para un gran número de avances, no es imposible sostener el anterior punto de vista, pero resulta que fueron precisos otros elementos/desarrollos para que se abriese realmente una nueva era científica, tecnológica y social. Así, sucede que aunque a finales de la década de 1960 la biología molecular podía presumir de tener tras de sí ya casi dos décadas de tradición, en las que se habían producido avances notables (había sido posible, por ejemplo, describir con gran detalle la maquinaria celular responsable de la replicación del ADN, la síntesis de proteínas, determinar en términos moleculares los principales mecanismos de transmisión de información genética en bacterias, o explicar el comportamiento genético de las formas más sencillas de vida, como las células procariotas, que no poseen núcleo), un rasgo llamativo es que no había conducido a ninguna aplicación práctica importante. Un informe emitido en 1975 por una compañía de ingeniería genética (Cetus Corporation) afirmaba: «Las aplicaciones prácticas de la biología molecular han sido lentas en llegar. En el nivel aplicado, todavía en este momento es difícil encontrar alguna habilidad médica o industrial realmente importante para la que sea esencial que conozcamos el código genético o que el ADN aparezca en la naturaleza como una doble hélice». El problema —o parte importante del problema— es que no se conocían técnicas adecuadas para manipular el ADN en el tubo de ensayo. Debido a que las moléculas de ADN son de gran tamaño, cuando se las intentaba fragmentar los cortes se producían al azar, con lo cual se descomponía la información genética contenida en ellas de forma tal que era prácticamente imposible de recomponer.

Biólogos moleculares en una escalera de espiral. De abajo a arriba: Christian de Duve, Francis Crick, James Watson, Jean Dausset, Thomas Cech, Werner Arber, Paul Berg, François Jacob, Renato Dulbecco, Manfred Eigen y Giorgio Bernardi.

«El Club de la corbata de ADN»: de izda. a dcha., Francis Crick, Alexander Rich, Leslie Orgel y James Watson (Cambridge, otoño de 1955).
Fue a finales de la década de 1960 y comienzos de la de 1970 cuando comenzaron a ser desarrolladas las técnicas necesarias para que la biología molecular «saliese al exterior», para que su hogar dejase de ser únicamente el laboratorio dedicado a «la búsqueda del saber por sí mismo». Con tales técnicas pudo hablarse ya con seguridad y realismo de términos como «biotecnología» o «ingeniería genética» (un término este último acuñado en 1965 por Rollin D. Hotchkiss [n. 1911]), la disciplina que se ocupa de «unir genes»; esto es, de sustituir un segmento de ADN de una célula por uno de otra (al organismo que surge de este proceso se le denomina transgénico). Por aquellos años, en efecto, se encontraron herramientas moleculares que podían resolver muchos de esos problemas: aislamiento (Bernard Weiss y Charles C. Richardson; 1967) de la enzima ligasa que puede unir cadenas de ADN; hallazgo (Linn y Werner Arber; 1968) de las denominadas enzimas de restricción, que son capaces de cortar por sitios determinados las moléculas de ADN. Disponiendo de instrumentos como estos, fue hasta cierto punto natural que en 1972 Janet Mertz y Ron Davis, de la Universidad de Stanford, descubriesen que se podían unir muy fácilmente moléculas de ADN cuando se las cortaba utilizando la enzima de restricción EcoRI. Las investigaciones de Mertz y Davis determinaron que al año siguiente Herb Boyer y Stanley Cohen desarrollasen un método para reordenar moléculas de ADN en un tubo de ensayo con el fin de crear moléculas híbridas procedentes en parte de, por ejemplo, ADN de una mosca y en parte de ADN bacterial. Entonces sí, ya se abrió una nueva era, socio-tecnocientífica; una era a la que se le pueden poner varios nombres: la del ADN recombinante, la de la biotecnología, la de la ingeniería genética, o quizá de modo más general y probablemente más apropiado, a tenor de las consecuencias-aplicaciones que se vislumbran y favorecen, la de las ciencias biomédicas moleculares.
Nuevos mundos, científicos, industriales y morales
Como en otras ocasiones (recordemos el ejemplo de la fisión nuclear, y numerosos descubrimientos químicos, como el de la dinamita o el DDT), el nuevo mundo científico que se fue abriendo a partir de 1953 ha terminado conduciendo a todo tipo de situaciones, científicas al igual que sociales (morales o éticas, por ejemplo); situaciones a las que tenemos que enfrentarnos. Y no siempre es fácil.
La lista de esas nuevas posibilidades y situaciones se encuentra al cabo de la calle y aumenta constantemente; a veces uno se siente tentado a decir: aumenta cada día. La creación de nueva vida transgénica fue una de las posibilidades que más pronto comenzó a hacerse realidad. Un momento importante en este sentido fue cuando, en 1971, Janet Mertz presentó, en una reunión celebrada en el famoso Laboratorio de Cold Spring Harbor, en Long Island, un proyecto de experimento en el que se pretendía insertar el cromosoma de SV40 en una cepa de laboratorio de Escherichia coli, proyecto que condujo al mencionado resultado de Mertz y Davis de 1972. La perspectiva de que una bacteria —especialmente una que aparece de manera natural en el intestino humano— pudiese ser tratada para transportar el ADN de un virus capaz, bajo ciertas condiciones experimentales, de provocar tumores en roedores alarmó a algunos biólogos moleculares, con la consecuencia de que el experimento con el SV40 fue pospuesto.
Se abrió entonces un debate dentro de la propia comunidad científica acerca de si era razonable introducir restricciones en la tradicional libertad de investigación científica.
El presidente de la National Academy of Sciences estadounidense, Philip Handler, se inclinó por mantener el debate dentro de la comunidad científica, pidiendo a Paul Berg (n. 1926), director del Departamento de Bioquímica de la Medical School de la Universidad de Stanford (al que también pertenecía Mertz), que estableciese un comité para examinar el problema y proponer acciones a corto y largo plazo. Berg, que contemplaba el comité como un grupo informal (Handler, no obstante, afirmó más tarde que actuaba en representación de la Academia), cumplió con el cometido, reuniendo, en abril de 1974, a un grupo de diez destacados biólogos moleculares y bioquímicos (entre ellos se encontraban David Baltimore, James Watson, Sherman Weissman, Daniel Nathans y Norton Zinder).
El producto de la reunión fue una carta, publicada en julio de 1974 en los Proceedings of the National Academy of Sciences, Science («Potential biohazards of recombinant DNA molecules» [«Riesgos biológicos potenciales de las moléculas de ADN recombinante»]) y en Nature («NAS ban on plasmid engineering» [«La Academia Nacional de Ciencias prohíbe la ingeniería plásmida»]).
«En una iniciativa sin precedentes —se comentaba en Nature—, la Academia Nacional de Ciencias ha solicitado que se establezca una moratoria mundial voluntaria en un área de la investigación científica debido a riesgos potenciales e impredecibles para la salud humana.» Específicamente, se pedía que se detuviesen dos tipos de experimentos hasta que se hubiesen evaluado mejor sus riesgos, o desarrollado métodos para impedir su propagación (uno de ellos era la unión de ADN de virus oncogénicos de animales, o de otros tipos, a elementos de ADN que se reprodujesen de manera autónoma). La carta del «Comité Berg» proponía también que se celebrase una conferencia internacional «para pasar revista a los progresos científicos en esta área». Ésta se celebró en el Centro de Conferencias de Asilomar, en Pacific Grove, California, en febrero de 1975. Participaron 150 científicos. David Baltimore, del Centro de Investigación del Cáncer del Instituto Tecnológico de Massachusetts (MIT) y que unos meses más tarde sería galardonado con el premio Nobel, inauguró la conferencia. Su intervención dejó claro que lo que se pretendía analizar eran únicamente los riesgos asociados a la investigación genética y las medidas de seguridad que se podían introducir. Específicamente, descartaba como temas «periféricos a esta reunión», «la utilización de esta tecnología en la terapia génica o ingeniería genética, que nos conduce a cuestiones como lo que está bien y lo que está mal, complicadas cuestiones de motivación política, y de las cuales no creo que este sea el momento adecuado [para discutirlas]».
Naturalmente no todo el mundo estaba de acuerdo con semejante planteamiento, pero yo no quiero entrar en esta cuestión aquí, ni comentar el hecho de que la conclusión de la conferencia fuese la de recomendar que se levantase la moratoria parcial impuesta en julio, reemplazándola con unas normas para la investigación en ingeniería genética (muchas de las cuales, por cierto, eran bastante estrictas). Lo que me interesa es mostrar que estaba claro, y todos lo apreciaron, comenzando por los científicos, que se había abierto un mundo cuyas implicaciones no se detenían en el laboratorio. Un mundo que incluye no sólo cuestiones relativas a la seguridad de los experimentos que se realizan, o la creación de posibles nuevas formas de vida, sino también otras más sociopolíticas y económicas. Como la espinosa cuestión de la posibilidad de patentar las variedades creadas de organismos vivos (microorganismos, plantas o animales transgénicos). En 1987, la Oficina de Patentes estadounidense concedió el derecho a patentar animales domésticos transgénicos, esto es, creados mediante ingeniería genética. Utilizando este acuerdo, el 12 de abril de 1988 se aceptaba la patente de un ratón transgénico producido en la Universidad de Harvard. Antes, en 1987, la misma Oficina había prohibido que se patentasen seres humanos alterados genéticamente, basándose en una enmienda antiesclavista de la Constitución norteamericana que impide la propiedad de seres humanos. Pero existe un amplio territorio intermedio: vivimos, lo sabemos demasiado bien, entre luces y sombras. Myriad Genetics, la compañía biotecnológica estadounidense que anunció el descubrimiento del primero de los genes asociados al cáncer de pecho hereditario, conocido como BRCA1 (estrictamente, la mutación de este gen sólo está relacionada con una pequeña fracción de esos cánceres), mostró con rapidez su intención de patentar el hallazgo, abriendo de esta manera el debate acerca de si se deben conceder patentes sobre genes individuales.
Sabemos ya también, gracias a toda esta ciencia biológica, que pequeños cambios en la estructura química de los genes pueden tener consecuencias muy importantes. En 1986, por ejemplo, un equipo de investigadores estadounidenses identificó el defecto genético responsable de un tipo de distrofia muscular. En 1989, un grupo de biólogos anunció el descubrimiento de la situación del gen que, cuando sale defectuoso, produce la fibrosis quística, una enfermedad que afecta a los pulmones, páncreas y otros órganos. En 1993, se localizó el gen de la corea de Huntington, un trastorno que produce una degeneración progresiva del cerebro, acompañada de la aparición de fuertes movimientos incontrolados y que conduce, inevitablemente por el momento, a la muerte. Y la lista crece casi cada día. Entre los trastornos de origen génico parece que se encuentran algunos como cánceres de colon, páncreas, pulmón y mama, diabetes, epilepsia, enfermedad de Tay-Sachs, esclerosis lateral amiotrófica (o enfermedad de Lou Gehrig), hemofilia, hipertensión arterial, neurofibromatosis y diversos síndromes (como el de Down).
Conocimientos como estos son buenos, ¿cómo no van a ser buenos? Pero pueden utilizarse también en direcciones que creo, o me hago la ilusión, no serían tan bienvenidas por los que aman la justicia. Las compañías de seguros al seleccionar a sus asegurados, o, simplemente, empresas al contratar a sus empleados, podrían utilizar el conocimiento de genomas individuales para introducir marginaciones inadmisibles, una situación esta cada vez más peligrosa, toda vez que el conocimiento del genoma humano camina a pasos agigantados. En efecto, el 26 de junio de 2000 la empresa privada estadounidense Celera Genomics, dirigida por el emprendedor científico Craig Venter (n. 1946), y la organización pública Proyecto Genoma Humano, en el que participaban científicos de, especialmente, Estados Unidos y el Reino Unido, al igual que de Francia, Alemania, Japón y China, efectuaron un primer anuncio conjunto manifestando que habían completado la secuenciación (la determinación de la estructura, de la «geografía») del genoma humano. A pesar de lo grandilocuente de aquella declaración, en la que estuvieron presentes el presidente Bill Clinton y el primer ministro británico, Tony Blair, todavía quedaba —queda— mucho que hacer. No se había dicho nada, por ejemplo, de cuántos genes forman el genoma humano; a comienzos de 2001 se superó tal carencia, señalándose que existen unos 30.000, frente a la cifra de alrededor de 100.000 que habían manejado hasta entonces los científicos (recientemente la cifra se ha reducido más, a 23.000). Y todavía subsisten cuestiones fundamentales: ¿a quién, a qué persona, de qué grupo, corresponde el genoma anunciado? A nadie estrictamente. El catálogo final de la secuencia de ADN humano es un mosaico de una persona hipotética, algo así como la interpolación de muchos genomas individuales. Semejante información es, por supuesto, esencial para numerosos fines, pero si de lo que se trata es de abordar problemas sanitarios individuales, lo que se necesita es ser capaz de secuenciar con cierta rapidez el genoma de personas concretas. Y esto es algo que ya se está haciendo: Craig Venter es uno de los primeros (si no el primero) que ha determinado la estructura de su genoma, como ha comentado en su autobiografía, A Life Decoded (Una vida descodificada; 2007).
Otro tema de nuestro tiempo que ha hecho posible la tecnociencia biológica es el de la clonación. El 25 de noviembre de 1996, Nature recibía un escueto artículo titulado «Viable offspring derived from fetal and adult mammalian cells» («Descendencia viable derivada de células fetales y adultas de mamífero»), que fue publicado en el número del 27 de febrero del año siguiente. Sus autores eran Ian Wilmut, el líder del grupo, A. E. Schnieke, J. McWhir, A. J. Kind y K. H. S. Campbell, del Roslin Institute de Edimburgo. No se trataba de un artículo cualquiera, uno más de los miles que constantemente irrumpen en la literatura científica, sino de uno en el que se anunciaba que se había conseguido traspasar una frontera muy especial: la reproducción de un mamífero, una oveja —Dolly— transfiriendo la carga genética (el ADN) del núcleo de una célula de glándula mamaria —es decir, una célula somática, no germinal— de una oveja adulta a un óvulo no fecundado y enucleado. Se había, en suma, clonado un mamífero.
Es cierto que Dolly no fue el primer clon de un mamífero; estrictamente, los gemelos son clones, pero lo son de un tipo muy diferente: se trata de una clonación natural, y más absoluta, por cierto, ya que al surgir de una escisión embrionaria, poseen no sólo idéntico ADN, sino también el mismo citoplasma (la sustancia que rodea al núcleo en donde se encuentra el ADN), mientras que clones como Dolly sólo tienen el mismo ADN que la célula que aporta la carga genética (en la medida que el ADN no opera aislado, sino que se halla en diálogo constante con su entorno citoplasmático, Dolly no es, por tanto, un verdadero clon, sino un clon del ADN, o «clon genómico»). Antes, en agosto de 1995, el grupo que había creado a Dolly produjo dos ovejas clónicas de raza Welsh Mountain —Megan y Morag—; la diferencia con Dolly es que procedían de células que, recogidas en embriones de nueve días, fueron cultivadas, diferenciándose en el cultivo: habían sido clonadas a partir de células embrionarias, no adultas.

Ian Wilmut con la oveja clónica Dolly.

Portadas de los números de Nature (15 de febrero de 2001) y Science (16 de febrero de 2001) en los que se presentaba la secuenciación del genoma humano.

Poco después de estos acontecimientos, en 1997, fruto también de los trabajos del grupo escocés, nacía Polly, con la que se dio una vuelta más al tornillo de la reproducción clónica, mostrando lo que con gran probabilidad vendrá en el futuro: su clonación tuvo lugar a partir de células cultivadas y transformadas genéticamente (se les añadió un gen humano en el proceso).
Las consecuencias de todo esto —sin olvidar las posibilidades que están abriendo últimamente las denominadas «células madre»; esto es, células que no han perdido la capacidad (antes supuesta únicamente en los primeros estadios de la vida) de generar todo tipo de tejidos— son inmensas y radicales. «Con el paso de las décadas y de los siglos —han manifestado Ian Wilmut y Keith Campbell en un libro divulgativo (significativamente titulado: La segunda creación)—, la ciencia de la clonación y las tecnologías que de ésta se deriven afectarán a todos los aspectos de la vida humana; el tipo de cosas que la gente pueda hacer, la forma de vivir e incluso, si lo deseamos, el tipo de personas que seremos.»
Es escalofriante escuchar opiniones como estas. Opiniones, además, muy verosímiles. Al oírlas, por un lado nos embarga una sensación de ilusión ante el futuro, si no para nosotros mismos, al menos para nuestros hijos, cuyas vidas pueden ser, en numerosos aspectos, más seguras y felices. Pero por otro, el temor nos invade: sabemos demasiado bien de los excesos de que son —han sido, cuando menos— capaces los humanos (los sufrimientos que generaron las creencias eugénicas en el siglo XIX y primeras décadas del XX vienen enseguida al recuerdo).
No es, por supuesto, la primera vez que nos enfrentamos a este tipo de situación. Al fin y al cabo, en más de un sentido, la ciencia no es sino un instrumento —un magnífico instrumento, cierto es— que necesita, ineludiblemente, de los humanos (de científicos al igual que de otros que no lo son) para poner de manifiesto todas sus potencialidades. Y éstos, los humanos, son, somos, capaces de lo mejor al igual que de lo peor. Es el problema de la ambivalencia de la ciencia, una característica de esta fuente de conocimiento a la que muchos aluden con frecuencia, con la intención —parece evidente— de socavar su estatus e implantación social. No niego, no puedo negar, cuán cierto es que el conocimiento que suministra la ciencia ha sido, es, y me temo seguirá siendo, origen de abusos y sostén de desigualdades que no conducen, precisamente, a facilitar lo que constituye su fin último: erradicar la ignorancia y el desamparo. Sin embargo, piénsese en lo que sería, hoy, la vida material e intelectual de los humanos (no de todos, lo sé, pero sí de muchísimos) sin lo que la ciencia les ha dado y da. Seríamos más ignorantes, más indefensos (al menos frente a la naturaleza y las enfermedades, seguramente también frente a otros humanos); seríamos, en definitiva, mucho más pobres en todos los sentidos. Por eso, por todos los bienes que hemos recibido de la ciencia, no deberíamos mirarla con recelo en una época como la presente, en la que de su mano se esperan profundas transformaciones que tienen como objeto preferente la propia vida. Debemos, eso sí, ser especialmente cuidadosos, estar particularmente atentos, al uso que se hace y hará de los nuevos conocimientos que se avecinan: la ciencia, no lo olvidemos, no está por encima de los humanos, no es un nuevo Dios al que debemos someternos, obedientes y sumisos. Para ello, para cumplir con esa tarea de vigilancia, no es preciso —no debería ser preciso— más que recurrir a los mecanismos de control de que se han provisto las sociedades democráticas. Pero no hay mecanismo posible de control, es imposible ejercer plenamente cualquier derecho, desde la ignorancia y el temor. Lo que yo he pretendido en las páginas precedentes es ayudar a subsanar algo de esa, ¡ay!, tan extendida ignorancia; contribuir a que se pueda comprender qué es la ciencia, algunos de sus contenidos y personajes, al igual que algunas de sus características. Evidentemente, lo he hecho desde la esperanza y la confianza para con ella, y no desde el temor o la suspicacia.