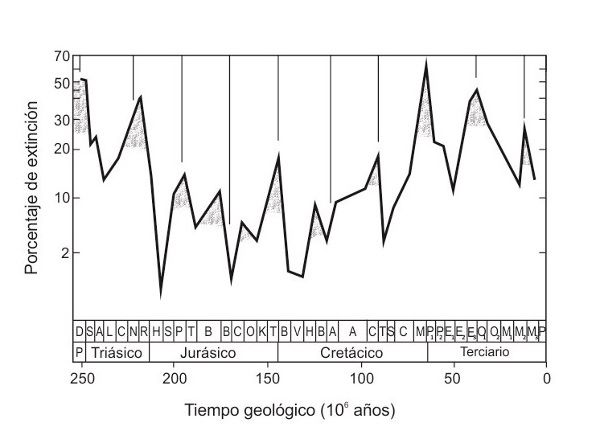
FIGURA 4.
La historia de la vida está puntuada por extinciones en masa. Los picos del gráfico indican periodos de extinción elevada; a la izquierda, se muestra el porcentaje de familias extintas.
Cortesía de David Raup y John Sepkoski.
La vida en la Tierra tiene más de 3800 millones de años, pero sólo los organismos formados por muchas clases diferentes de células parecen fascinar a los apasionados por el diseño. Después de que tales organismos evolucionaran, hace 600 millones de años, el infierno se desató y la historia de la vida en la Tierra ha seguido desde entonces una pauta compleja.
En los pocos millones de años que siguieron a ese hito capital en la historia terrestre, los mares se llenaron de miríadas de formas de vida, seres nadadores, merodeadores, sedentarios, así como excavadores. El acontecimiento fue tan espectacular que el término coloquial de explosión cámbrica no es una exageración. Tres mil millones de años de abrumadora simplicidad biológica se vieron sustituidos de la noche a la mañana —desde una perspectiva geológica— por una efervescente complejidad.
Una vez establecida, la vida multicelular siguió una ruta ascendente en la escala de complejidad, de modo que los mares modernos contienen el doble de especies que los del mundo cámbrico. Sin embargo, este incremento no fue una tendencia estable, donde cada era se apuntaba rutinaria y predeciblemente nuevas ganancias en la diversidad. El transcurso del tiempo estuvo marcado por una renovación continua, donde nuevas especies sustituían a las existentes. Y, lo más espectacular, toda progresión regular desde los tiempos antiguos a los modernos se vio interrumpida por catastróficos colapsos ocasionales en la diversidad, extinciones en masa que, en un caso, afectaron al 96 por ciento de las especies existentes en un instante geológico.
Cinco episodios de este tipo puntúan la historia de la vida. También infligieron pérdidas muchos colapsos menores, que no son lo suficientemente grandes como para merecer la denominación de «extinción masiva», pero que sin embargo fueron devastadores a escala continental. El resultado es que el 99,9 por ciento de todas las especies que vivieron alguna vez están hoy extinguidas. En palabras de un estadístico bromista: «Según una primera aproximación, todas las especies están extinguidas». A pesar de nuestra insignificancia estadística, somos, junto con las demás especies existentes —entre 10 y 30 millones—, la última expresión de un proceso de 600 millones de años de creación y extinción.
Dos pautas principales dominan esta historia. La primera es su principio, la explosión cámbrica, única en diversos aspectos importantes. La segunda es el repetido colapso de la biodiversidad, las extinciones en masa y sus primos menores. Para que la nueva ciencia de la complejidad —con sus nociones duales de autoorganización y límite del caos— tenga interés para los biólogos, deber ser capaz de iluminar de algún modo directo o, incluso, indirecto esas dos pautas principales. En el extenso paisaje de la historia de la Tierra estamos buscando las huellas de la complejidad, por débiles que sean.
***
«Hola, Roger, creo que tengo la respuesta», dijo una voz exultante al otro lado de la línea telefónica. «Creo que puedo explicar la pauta. Tiene que ver con mis relieves rugosos». Esto ocurrió a finales de julio de 1988, y quien llamaba era Stu Kauffman. Ese mismo mes había publicado un artículo en Science titulado «A Lopsided Look at Evolution», en el que describía algunas de las preguntas planteadas por la explosión cámbrica y las recientes ideas acerca de su solución. «Los relieves rugosos explican por qué hay innovación de alto nivel en el Cámbrico, pero no más tarde», continuó Stu.
La pregunta clave sobre la naturaleza de la explosión cámbrica está relacionada con la innovación evolutiva, no tanto con su cantidad, que fue grande, como con su calidad, que fue extraordinaria. «Sin precedentes y nunca superada», así me la describió James Valentine. Valentine, que en la época del artículo de Science estaba en la Universidad de California, Santa Barbara, pero que ahora trabaja en Berkeley, ha hecho un extenso estudio de la explosión cámbrica y sus consecuencias. «Es el fenómeno único más espectacular del registro fósil», dijo.
Es cierto que ha habido con posterioridad enormes estallidos de innovación en la historia de la vida, la mayoría tras las extinciones en masa. Por ejemplo, después de la extinción pérmica hace unos 250 millones de años, en la que se estima que pereció el 96 por ciento de las especies existentes, el ritmo de innovación alcanzó casi el del Cámbrico. Pero la innovación fue debida principalmente a variaciones sobre los temas existentes; no se añadieron nuevos temas principales. En el Cámbrico, en cambio, la innovación se produjo en gran medida en el nivel de la producción de nuevos temas, con un número relativamente menor de variaciones sobre ellos. «Éste es el reto», dijo Valentine. «Hay que explicar el paso de pocas especies en muchos grupos durante el Cámbrico a muchas especies y menos grupos más tarde».
Los grupos de los que Valentine estaba hablando se encuentran en los niveles superiores de la estructura jerárquica de la clasificación biológica: las clases y, en particular, los tipos. Los tipos, justo por debajo del nivel del reino (animales y plantas, en la clasificación tradicional, «de sentido común») y justo por encima de las clases (mamíferos, reptiles, etcétera), representan diseños corporales principales en la diversidad de la vida. Por ejemplo, los artrópodos, el más poblado de todos los tipos, tienen apéndices articulados, e incluyen criaturas como los insectos, los ciempiés, los miriápodos, las arañas y los cangrejos. Los humanos, y todos los demás vertebrados que tanto dominan nuestra visión del mundo, forman parte del tipo de los cordados. Los tipos son diseños corporales discretos, sobre los que pueden crearse muchas variaciones.
Hay en el mundo actual treinta tipos principales, más o menos como en los últimos 500 millones de años, una sorprendente continuidad de los diseños anatómicos, a partir de los cuales han existido y desaparecido hasta 50 000 millones de variantes. Tras la explosión cámbrica, pudo haber hasta un centenar de tipos, la mayoría de los cuales se extinguió enseguida, con lo que se llegó al nivel moderno de diversidad. «Tanta innovación no fue exclusiva del Cámbrico, pero sí lo fue su magnitud», dijo Valentine, planteando el problema de forma sucinta. Lo que el mundo cámbrico y poscámbrico inmediato experimentó fue una tremenda experimentación evolutiva, seguida de un severo proceso de selección.
El descubrimiento de esta pauta es una de las grandes historias de la historia de la paleontología. Empieza básicamente con Charles Darwin, finaliza con Harry Whittington, catedrático de geología en la Universidad de Cambridge, y ha sido narrada por Stephen Jay Gould en su libro La vida maravillosa. Un actor clave en esta historia fue Charles Walcott, secretario del Instituto Smithsoniano y descubridor en 1909 de la más espectacular ventana al mundo cámbrico, Burgess Shale. Los depósitos de Burgess Shale, que contienen una sorprendente encapsulación de vida costera durante un breve momento de la historia cámbrica, están situados en las montañas Rocosas de la Columbia Británica, Canadá. Para Walcott, la vista a través de esa ventana era la de un mundo con las mismas pautas que hoy, más primitivo, sin duda, pero con igual diversidad. Estaba siendo un buen darwinista.
Como Darwin concebía la selección natural como un proceso esencialmente gradual, capaz de producir gran innovación, pero sólo a partir de pequeños incrementos en el transcurso de largos periodos de tiempo, puso reparos a la idea de un cambio rápido. Darwin consideraba que el cambio rápido, ya fuera en la aparición o en la desaparición de especies, era un desafío a su teoría. Explicó la aparentemente abrupta emergencia de vida multicelular en el Cámbrico diciendo que esas formas anteriores debían de haber existido, pero estaban ocultas por la imperfección del registro fósil. «[D]urante esos vastos y todavía desconocidos periodos de tiempo, los seres vivientes hormigueaban en el mundo», escribió sobre el periodo precámbrico en El origen de las especies.
Cuando Walcott descubrió la profusión de formas de vida de Burgess Shale, que duplicaba lo que se sabía hasta entonces del Cámbrico, se agudizó en gran medida el desafío de la aparición rápida. Los precursores de los animales del Cámbrico, cuya existencia postuló Darwin, no fueron descubiertos en las cantidades predichas. Y la abundancia de la vida cámbrica, tan aparente en Burgess Shale, hizo aún más explosivo el instante de aparición rápida. Walcott, un darwinista ferviente, tenía dos elecciones.
O bien podía decir que el descubrimiento de Burgess Shale confirmaba una aparición explosiva de vida en el Cámbrico y demostrar así que Darwin se equivocaba, o bien considerar las criaturas de Burgess en el contexto del gradualismo y la continuidad —negando la aparición súbita de una gran diversidad— y mantener de ese modo el punto de vista darwinista. Eligió lo segundo. Para Walcott, y para las dos generaciones que lo siguieron, el Cámbrico no sólo era el producto de una larga historia de evolución, sino que también presagiaba a la perfección el mundo presente, con sus treinta tipos, más o menos. Al asignar a los tipos modernos prácticamente todas las criaturas que encontró en los depósitos, por extrañas que fueran, Walcott cerró los ojos a la diversidad real que estalló en la vida durante el Cámbrico.
Hasta la llegada de Harry Whittington y sus estudiantes en las décadas de 1960 y 1970 no se reconoció la naturaleza verdaderamente inusual de gran parte de la fauna de Burgess. Whittington, experto en trilobites, descubrió que Walcott había asignado muchas criaturas cámbricas a grupos que no les correspondían. Al final, se propusieron un centenar de tipos, no los treinta supuestos por Walcott y que todos habían creído. No cabía duda entonces de que el Cámbrico tenía que concebirse como un acontecimiento de innovación evolutiva explosiva. Y, también, que se había visto seguido de una extinción masiva, que moldeó el mundo que conocemos.
***
«Hay dos hipótesis rivales para explicar la pauta de innovación en el Cámbrico», me dijo Valentine. «La primera es la hipótesis ecológica, por la que tengo cierta inclinación, y la segunda es la teoría genómica. Quizá no sean del todo rivales», añadió. «Quizás haya un poco de cada». La teoría ecológica es sorprendente por su sencillez. Los primeros organismos cámbricos entraron en un mundo desprovisto de competidores, un mundo que debió de rebosar de bacterias y algas unicelulares, que representaba más una fuente potencial de alimento que un reto competitivo. Dada la cantidad de nichos ecológicos disponibles, fueron viables todas las clases de variantes evolutivas: la evolución procedió por lo tanto a través de grandes saltos, más que por medio de pequeños pasos graduales. «Después de la extinción pérmica, aun cuando se habían perdido innumerables especies, toda la gama de nichos ecológicos habría seguido estando ocupada», afirmó Valentine. «Las oportunidades ya no estaban ahí, como había ocurrido en el Cámbrico». La innovación en el pospérmico, por lo tanto, fue mucho más restringida, o eso es lo que afirma la hipótesis ecológica.
Según la hipótesis genómica, la evolución produjo en el Cámbrico tantas formas experimentales porque las dotaciones genéticas de las especies —los genomas— carecían del grado de coherencia y el estrecho control que desarrollaron más adelante. Por ello fue posible y viable una mutación más drástica, con la consiguiente generación de temas en lugar de variaciones sobre ellos.
La hipótesis ecológica está animada esencialmente por factores externos, a saber, la oportunidad ecológica; la hipótesis genómica, por factores internos, la viabilidad de la mutación sustancial. «Ninguna de las dos es correcta», proclamó Stu Kauffman, en ese día de julio de 1988. «Aunque la solución tiene más que ver con los genes que con la oportunidad ecológica».
Stu había estado trabajando en su noción de relieves adaptativos rugosos, pero no había llegado a las ideas sobre relieves coevolutivos y el límite del caos. Eso vino más tarde. Las mutaciones que afectan al desarrollo embrionario al principio del proceso pueden producir una alteración espectacular en la forma adulta, porque el pequeño cambio inicial se ve aumentado a medida que tiene lugar el desarrollo, explicó Stu. «El desarrollo embrionario representa un relieve adaptativo rugoso, en el que se llega muy pronto a los óptimos locales». ¿Lo que quiere decir que alcanzar variantes más aptas al principio del desarrollo se hace cada vez más difícil?, insinué. «Eso es. La ontogenia temprana se queda congelada, y las nuevas variantes tienen que venir cada vez más del desarrollo posterior, lo cual provoca cambios evolutivos menos drásticos. Los seres cámbricos pudieron explotar nuevas variantes de eficacia biológica al principio de la ontogenia y dar grandes saltos evolutivos, pero cuando llegó el Pérmico ese juego ya se acabó y sólo fueron posibles pequeños cambios». Stu escribió sus ideas, las publicó en la revista Evolutionary Ecology y me envió un ejemplar. No pensé más en ello. Hasta que apareció la complejidad.
Pregunté a Stu recientemente si pensaba que la explicación del paisaje rugoso seguía siendo válida. «Sí», contestó. «Pero creo que habría que considerarla en un contexto coevolutivo, cosa que no hice antes». En ese caso, dije, ¿cómo aplicarías aquí tu modelo coevolutivo del límite del caos? Se produjo una pausa, poco característica y lo suficientemente prolongada como para darme cuenta de que entrábamos en territorio inexplorado. Yo había pensado la explosión cámbrica en términos cualitativos, en su forma más simple, un salto de los organismos unicelulares a los multicelulares. Había pensado la clase de interacciones ecológicas que debieron de desarrollarse entre poblaciones de organismos en el mundo precámbrico, poblaciones mezcladas de bacterias y algas en los estromatolitos, por ejemplo. Y, en un sentido abstracto, esas miniecologías complejas se convirtieron en interacciones al alcance de un único organismo en el mundo poscámbrico: se había producido un salto jerárquico. Pero ¿y el límite del caos?
¿Resulta razonable pensar en los animales cámbricos como parte de un sistema en régimen caótico, un sistema moviéndose hacia el límite del caos, pero sin haberlo alcanzado todavía?, pregunté a Stu. «Eso tendría sentido», dijo. «En mi modelo tendrían que estar interaccionando entre sí de forma muy estrecha. Si hubieran evolucionado recientemente a partir de unos pocos antepasados comunes, eso sería razonable». Si la explosión cámbrica significó de verdad un régimen caótico, las perturbaciones causaron grandes avalanchas, según el modelo de Stu. Y eso incluiría una mayor propensión a la innovación evolutiva, lo cual habría podido producir novedades inusualmente innovadoras. A medida que el sistema coevolucionó hacia un estado de equilibrio (el límite del caos, en el modelo de coevolución de Stu), disminuyeron las respuestas a las perturbaciones, la innovación se hizo menos arriesgada, hasta que se alcanzó un estado de renovación estable. «Todo lo que se necesita para explicar la diferencia de innovación tras la extinción pérmica es que el sistema sea empujado de nuevo hacia el régimen caótico, pero no demasiado», afirmó Stu. «Habría innovación en el rebrote postextinción, pero sería menos exagerada».
Plausible, pero era una completa especulación. ¿Cómo podríamos saberlo?, pregunté. ¿Qué pista podríamos buscar? Esta vez no hubo vacilación. «Hay que mirar las extinciones», fue la respuesta instantánea de Stu. «Si esta disparatada idea tiene algún valor, los índices de extinción en el Cámbrico también tienen que haber sido más elevados, y eso significaría que las vidas de las especies habrían sido más cortas». Y en otros periodos después de extinciones en masa, añadí. «Sí, exacto. ¿Qué pruebas tenemos?». Sentíamos que podíamos estar cerca de algo nuevo e interesante. No lo sé, pregunté. Lo averiguaré. ¿El resultado? Pregunté a paleontólogos que debían saberlo y, una y otra vez, obtuve la misma respuesta: no hay datos fiables.
***
Abandoné mi incursión en el proceso de innovación del Cámbrico y me enfrenté a otras dos preguntas. La primera se refiere a su pauta global de innovación. ¿Es una pauta única o hay ahí algo más general, algo que pueda iluminar de un modo fundamental el proceso de innovación en los sistemas complejos? La segunda pregunta plantea si los productos de ese proceso de innovación son únicos o no. Si pudiéramos hacer retroceder el reloj y repetir la explosión cámbrica, ¿sería el mundo como hoy? ¿Habría seres humanos para observarlo y meditar sobre él?
Cuando George Gumerman leyó un artículo sobre la fauna de Burgess Shale y la explosión cámbrica en el número del 20 de octubre de 1989 de Science, encontró suficientes semejanzas en la dinámica del sistema como para examinar de nuevo su propia área de interés: la evolución de las sociedades prehistóricas sudoccidentales. Como uno de los coorganizadores del congreso de Santa Fe sobre complejidad social en el Sudoeste, George había considerado pertinente la pauta de la innovación. Señaló que una gran diversificación en las convenciones sociales se produjo en el Sudoeste durante un siglo y medio, entre los años 1000 y 1150; y a ello siguió una reducción igual de drástica en la diversidad, con el dominio de la cultura del cañón del Chaco. Experimentación seguida de especialización; es la pauta de la vida en el Cámbrico y la de la vida de los anasazi en el sudoeste norteamericano. ¿Hay otros ejemplos?
«Se aprecia también en la innovación tecnológica de las sociedades industriales», dijo Stu a mi subgrupo en el congreso sobre el Sudoeste. «Pensad en las primeras bicicletas o los primeros coches», continuó. «Mucha experimentación al principio, diferentes formas de bicicleta, diferentes formas de propulsión y diseño de los coches, todas viables. Pasa el tiempo y el mundo se llena de bicicletas o coches, o de cualquier otra cosa, los extremos quedan suprimidos, sobreviven unas pocas formas y la consiguiente innovación se centra en la mejora de los temas que quedan. Se pasa de la generación de muchos temas a las variaciones sobre unos pocos, como en el Cámbrico».
Sin embargo, con la innovación humana siempre existe la posibilidad de dar vida otra vez a un diseño desaparecido. Podríamos, por motivos de moda o necesidad económica, resucitar el velocípedo de rueda alta o el automóvil de vapor.
Pero, para la historia evolutiva, las formas perdidas son sólo eso: historia. La extinción es para siempre, y la reevolución de grandes formas extintas exige la concatenación de demasiados acontecimientos improbables como para que se produzca.
Los únicos trilobites y dinosaurios con que nos encontraremos son los de los libros, las colecciones de los museos o las exposiciones geológicas. Así, la contingencia histórica influyó en la forma de la explosión cámbrica y su secuela. Los tipos que se perdieron, bien por exigencias de la competencia o por eliminación estocástica (en otras palabras, por la mala suerte), no se reinventaron nunca. La forma del mundo actual se vio, al parecer, influida en gran medida por los tipos que sobrevivieron hace 500 millones de años.
¿Y las tradiciones sociales y culturales que se extinguen tras periodos de experimentación, como ocurrió entre los anasazi hace casi un milenio? ¿Es su destino correr la misma suerte que los automóviles de vapor o los trilobites? «Puedes extender la pauta de la historia cámbrica a la historia cultural humana», me dijo George. «Hemos visto lo similar que es la pauta de innovación, con un estallido de novedades y luego la consiguiente pérdida. Y, de la misma manera que no vemos la reaparición de formas animales extintas, no vemos la reaparición de culturas una vez han cambiado». Hay tanto contenido histórico en las tradiciones culturales y las mitologías que, una vez perdidas, es improbable que se reformulen otra vez exactamente igual, explicó George. En otras palabras, las culturas se parecen más a los trilobites que a los automóviles.
¿Qué consistencia tiene, pues, una pauta de innovación en los sistemas complejos adaptativos como ésos? Cada uno de ellos se ve afectado por la contingencia histórica, pero según grados diferentes. Es sin duda significativo que, con todas esas diferencias de detalle —en los ámbitos biológicos, culturales y tecnológicos—, la pauta global sea notablemente similar. Ello alienta la creencia de que la consistencia de la pauta sea algo más que una simple coincidencia o una simple analogía. Puede estar en acción una dinámica fundamental, que haga predecible, hasta cierto punto, la pauta de innovación de los sistemas complejos adaptativos.
***
En su artículo de Science sobre la explosión cámbrica, Simon Conway Morris finalizaba con un pequeño experimento mental. «¿Y si la explosión cámbrica fuera a repetirse?», preguntó. Se produciría la misma innovación explosiva, aventuró, y, casi con seguridad, la misma reducción de la diversidad tras la innovación masiva. La fauna posterior ocuparía nichos instantáneamente reconocibles a nuestros ojos: herbívoros, carnívoros, insectívoros, etcétera… criaturas grandes y pequeñas, con vidas como las pasadas y las actuales. Pero, afirmaba, debido a la contingencia histórica, no se parecerían a nada de lo que hemos experimentado y serían «dignas de la más refinada ciencia ficción».
El hecho de la contingencia histórica, que Stephen Jay Gould ha defendido con más fuerza incluso en los últimos años, significa que el mundo en que vivimos es sencillamente uno de entre una infinidad de mundos virtuales. Si pasamos la cinta otra vez, afirma, incluso el más modesto recodo en la ruta de la historia se traducirá en un drástico efecto unos cien millones de años más tarde. Si se multiplican por un millón estas pequeñas excursiones del destino, el resultado final será un mundo irreconocible a nuestros ojos. ¿O no? ¿Existe una infinidad virtual de mundos posibles, de los cuales nuestra experiencia es sólo una? Según Brian Goodwin, no.
Para Brian, la mecánica del desarrollo embriológico está muy limitada, y eso restringe grandemente las clases de estructuras y las clases de especies que pueden surgir. En el lenguaje de los sistemas dinámicos complejos, el espacio de las posibilidades morfológicas está tenuemente poblado de atractores, esos estados en los que los sistemas dinámicos acaban por asentarse, especies fantasmas que podrían ser devueltas a la vida bajo las circunstancias correctas. Esta imagen es muy diferente de la visión convencional de la evolución darwinista, en la que los procesos de la selección natural y la adaptación pueden explorar prácticamente todos y cada uno de los rincones de ese espacio. Le pregunté a Brian si sostendría que en su mundo sólo era posible un número limitado de especies, mientras que en el mundo del adaptacionista se daban una infinidad de especies posibles. «Eso exagera un poco la cuestión», contestó Brian. «Incluso el más ferviente adaptacionista acepta algunas restricciones a las posibilidades morfológicas, como la biomecánica básica. Pocos sostendrían que los organismos terrestres podrían desarrollar ruedas, por ejemplo. Pero, en conjunto, tu comparación es correcta».
Para la mayoría de biólogos, el enfoque de Brian de los sistemas dinámicos es, en el mejor de los casos, apenas comprensible y, en el peor, del todo disparatado. «Es una conjetura plausible», me recordó otra vez Brian. De acuerdo, pero llevémosla un poco más lejos, respondí. ¿Afirmas que el mundo de ahí afuera está poblado por una gama de especies fantasma, atractores dinámicos, de las cuales sólo una parte puede ser ocupada en un momento dado? «Sí, es una afirmación correcta. Sin embargo, no habría que concebir los atractores como estáticos. Cambian, igual que las posibilidades dinámicas, al cambiar el entorno. En el pasado, he cometido el error de hacer caso omiso del entorno, pero es importante. Puede disminuir la estabilidad de algunos atractores y mejorar la estabilidad de otros».
Surgía una pregunta obvia acerca de la contingencia histórica. Si es cierto que sólo un número limitado de atractores pueblan el espacio morfológico potencial, aventuré, ¿significa eso que si repitieras la explosión cámbrica, el nuevo mundo no sería tan diferente del que conocemos? ¿Que Steve Gould podría no estar en lo cierto cuando dice: «El magnetófono divino contiene un millón de escenarios, cada uno perfectamente coherente»?
«Deja que te conteste a la pregunta de este modo», empezó reflexivamente Brian. «¿Eres consciente del fenómeno de la convergencia en biología, cuando ves una morfología sorprendentemente similar en especies muy divergentes?». ¿Como el lobo de Tasmania y el lobo de verdad?, insinué. «Sí, uno es un marsupial, el otro es un mamífero placentario, separados por, no sé, 50 millones de años. Y, sin embargo, en términos anatómicos son prácticamente iguales». La teoría evolutiva convencional explica el fenómeno en parte por la contingencia histórica: ambos descienden de un mismo antepasado mamífero. Pero el nudo del razonamiento es que las adaptaciones similares conforman anatomías y comportamientos similares. «Creo que eso es estirar un razonamiento débil hasta el punto de ruptura», dijo Brian. «No hay dos entornos tan similares como para producir una anatomía tan paralela».
¿Así que dirías que el lobo de Tasmania y el lobo de verdad son atractores en el espacio de las leyes morfogenéticas? «Sí». ¿Y extenderías el razonamiento a la repetición de la explosión cámbrica? «Lo extendería en parte», dijo Brian. «Pondré una analogía. Supón que repetimos la Gran Explosión. ¿Cuáles son las posibilidades de conseguir la misma tabla periódica de elementos naturales, las mismas noventa y dos combinaciones de protones, neutrones y electrones? Bastantes, o eso es lo que creo. Concibo una repetición de la explosión cámbrica del mismo modo, quizá no con la misma extensión, pero como una imagen. Si existen atractores dinámicos en el espacio de las posibilidades morfológicas, como creo, una repetición de la explosión cámbrica produciría un mundo mucho más parecido al que conocemos de lo que afirma Steve Gould. No sería idéntico, pero habría un montón de semejanzas, fantasmas que reconoceríamos en el acto».
Dicho de otro modo, la historia evolutiva quizá no sea una maldita cosa tras otra, sino, en una interesante medida, inevitable. Esto se está convirtiendo ya en una especie de estribillo de los sistemas complejos adaptativos.
***
«El estudio de las extinciones nunca ha estado de moda», se lamentó David Raup. «Hay muchas razones, estoy seguro. Pero es probable que pueda remontarse bastante a Darwin». Dave es geólogo en la Universidad de Chicago, donde el Departamento de Ciencias Geofísicas está situado en South Ellis, entre el falso gótico de gran parte de la arquitectura de la universidad. El departamento es, sin embargo, un edifico moderno, de ladrillo rojo y cristal, ventanas en ángulos ingeniosos y ese tipo de cosas. El despacho de Dave es grande, sobrio, iluminado por dos ventanas y, para ser el dominio de uno de los principales investigadores en ese campo, con una sorprendente ausencia de rocas y fósiles. A pesar de su severo aspecto exterior, el entorno investigador de Dave es más el ordenador y el análisis estadístico superior que la extracción de rocas de antiguos estratos. «Al estar comprometido con el gradualismo, Darwin intentó negar la existencia de extinciones en masa», explicó Dave.
Del mismo modo que intentó negar la abrupta aparición de los animales multicelulares en el Cámbrico, sugerí. «Sí, eso, Darwin dijo que la extinción era “el misterio más gratuito” o algo así. Pero también dijo que el origen de las especies es gradual, y también lo es su extinción». Una frase de El origen de las especies refleja esto: «Las especies y los grupos de especies desaparecen gradualmente, uno tras otro, primero de un lugar, luego de otro y, por último, del mundo».
Si las especies desaparecen como parte de la dinámica gradualista de la selección natural, ya no hay nada más que explicar. Ya no hay nada más que decir del proceso, excepto la necesidad de etiquetar como fracasos aquellas especies que se han extinguido. Darwin lo afirmó de modo explícito: «Los habitantes de cada periodo sucesivo de la historia del mundo han derrotado a su predecesores en la carrera de la vida y están por ello más arriba en la escala de la naturaleza». (Todo el mundo sabe que los dinosaurios son fracasos, ¿no es cierto?). La reticencia de Darwin a contemplar la desaparición abrupta de las especies se apoyaba no sólo en el modelo gradualista de la selección natural, sino también en la noción de gradualismo en la geología en general, que había sido promulgada por su amigo Charles Lyell.
A principios del siglo XIX, el barón George Cuvier, el gran geólogo francés, propuso lo que llegó a conocerse como teoría de las catástrofes o catastrofismo. Según esta teoría, los cambios abruptos en la fauna que los geólogos veían en los estratos de rocas eran resultado de devastaciones periódicas que borraban todas o casi todas las especies existentes, y el periodo siguiente era repoblado con nuevas clases de animales y plantas por la mano de Dios. Lyell rechazó semejante hipótesis no científica (como había hecho James Hutton antes que él) y la sustituyó por la noción de que los procesos geológicos procedían de forma gradual: todos los procesos geológicos. En su obra principal, Principios de geología, Lyell afirmaba que las transiciones abruptas del registro geológico se revelarían algún día como erróneas, cuando se descubrieran los estratos transicionales. El gradualismo acabó imponiéndose frente al catastrofismo. Las visiones del mundo de Darwin y Lyell eran, por consiguiente, perfectamente complementarias.
«Con el tiempo, los geólogos llegaron a aceptar los aparentes cambios abruptos del registro geológico como reales, como extinciones en masa, pero las explicaciones darwinistas —competencia, depredación, etcétera— subsistieron y dominaron el escaso debate que se produjo», dijo Dave. El debate intelectual en torno a los mecanismos de extinción siguió estando llamativamente ausente de la comunidad biológica. Entonces llegó Luis Alvarez. «Escandaloso en su comportamiento, escandaloso en sus afirmaciones, consiguió sin duda trastornarlo todo», dijo Dave con malicia. Alvarez, físico de la Universidad de California, Berkeley, afirmó en 1980, junto con varios colegas, que la extinción masiva ocurrida hace 65 millones de años y que provocó el fin de los dinosaurios, estuvo causada por la colisión de un asteroide gigante. A Dave le gustó la idea, en parte por su audacia, en parte por su plausibilidad. Cuatro años más tarde, Dave y su colega Jack Sepkoski dieron un paso —o varios— más allá que Alvarez y afirmaron que las colisiones de asteroides gigantes ocurrían cada 26 millones de años, provocando periódicas extinciones en masa.
Hoy en día, la mayoría de entendidos acepta que la extinción de fines del Cretácico fue causada por la colisión de un asteroide. La misma explicación se considera verosímil para unas pocas extinciones más. Pero la idea del impacto periódico, cada 26 millones de años, sigue siendo claramente polémica, como mínimo porque para la mayoría suena demasiado a la vieja idea del catastrofismo. Sin embargo, Dave ha seguido insistiendo y recientemente ha reunido las pruebas de la cronología de la colisión de asteroides (a partir de la edad de los grandes cráteres) y la cronología de las extinciones en masa (a partir del registro fósil) y ha demostrado que encajan bastante. «Hasta un 60 por ciento de todas las extinciones puede ser resultado de la colisión de un asteroide», concluyó. El sesenta por ciento. Eso es muchísimo, dije, sin ocultar mi incredulidad. «Lo sé. La gente no lo va a aceptar fácilmente, pero es una hipótesis creíble».
FIGURA 4.
La historia de la vida está puntuada por extinciones en masa. Los picos del gráfico indican periodos de extinción elevada; a la izquierda, se muestra el porcentaje de familias extintas.
Cortesía de David Raup y John Sepkoski.
***
En todas las causas propuestas para las extinciones en masa, incluyendo la colisión de asteroides, hay implícita una igualdad entre la causa y el efecto, entre la escala de la perturbación ambiental y la proporción de especies que desaparecen. Así es como funcionaría un mundo regido por ecuaciones lineales: grandes perturbaciones producen grandes extinciones; pequeñas perturbaciones, pequeñas extinciones. «Eso no es necesariamente cierto», dijo Stu Kauffman, «no si tenemos razón acerca de los ecosistemas en equilibrio en el límite del caos». El mundo que Stu estaba describiendo era un mundo no lineal, donde la dinámica compleja del tipo que ya hemos encontrado produce pautas complejas. «Puede ser que cambios similares en el medio ambiente produzcan extinciones de todas las magnitudes», añadió Stu.
Estábamos hablando en la Universidad de Pennsylvania; en el laboratorio vecino, bioquímicos de bata blanca manejaban preciosos líquidos con exquisita precisión, excursiones en la evolución molecular experimental, mientras en su despacho se discutían teorías sobre extinciones globales. Parecía extraño. ¿Estás hablando de tu sistema coevolutivo? «Sí», dijo Stu. «¿Recuerdas cuando nuestro ecosistema se puso él solo en un estado de equilibrio, el límite del caos, lo pellizcábamos un poco con alguna clase de cambio externo y producíamos avalanchas de cambio de todos los tamaños?». Me acordaba. ¿Como las avalanchas en el montón de arena de Per Bak, en equilibrio en el estado crítico, dando una distribución exponencial? «Exacto. Eso era parte de nuestro razonamiento de que el ecosistema informático había llegado al límite del caos».
¿Y lo que quieres saber es si los ecosistemas reales —ahí fuera— también están en equilibrio en el límite del caos? «Quizá la naturaleza ha hecho el experimento para damos la respuesta», respondió Stu. «Quizá la respuesta esté en los datos de las extinciones en masa». Stu y su colega Sonke Johnsen sacaron los datos sobre las extinciones en masa (de un artículo de Dave Raup), hicieron un gráfico con la magnitud de las extinciones y su frecuencia y buscaron una ley exponencial. «El resultado es muy parecido a una ley exponencial», dijo Stu. «No es una línea recta, es ligeramente convexa, pero así se comportan las extinciones en nuestro ecosistema informático». Si los datos de vuestro sistema coevolutivo son los mismos que los obtenidos con los datos de Dave a partir de extinciones reales, ¿significa eso que el mundo que vemos está en equilibrio en el límite del caos? «Así es como lo interpreto», dijo Stu. «Sólo que ligeramente en el lado congelado del límite del caos». Los ecosistemas, los informáticos y los reales, están justo en el lado congelado del caos, afirmó Stu, como resultado de una perturbación continua.
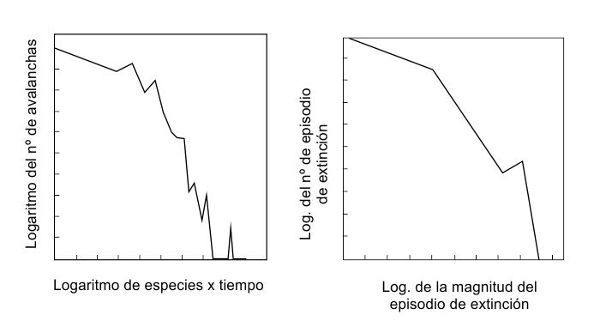
FIGURA 5.
Las extinciones en los ecosistemas informáticos y reales se acercan a una ley potencial. El gráfico de la izquierda muestra el logaritmo del número de avalanchas (equivalente a las extinciones) en el eje vertical y, en el eje horizontal, el logaritmo del tamaño de las avalanchas en el ecosistema informático de Kauffman, con 100 especies (ambos ejes están trazados como logaritmos naturales). Una ley potencial sería una línea recta descendente de izquierda a derecha. El gráfico de la derecha muestra lo mismo para las extinciones del registro fósil. Las pendientes son similares, y ambas están en el lado congelado del límite del caos.
«Si tenemos razón, no sólo sabemos algo de la dinámica de los ecosistemas en el mundo real en el contexto de los sistemas complejos adaptativos», continuó Stu, «sino que también nos enfrentamos a la noción contraintuitiva de que una extinción masiva como la de finales del Pérmico podría haber sido causada por el mismo tipo de perturbación que la producida por una pequeña oscilación en el mapa de las extinciones». Si tenéis razón. «Sí».
***
Dave sacudió lentamente la cabeza cuando le pregunté si pensaba que los datos de las extinciones en masa mostraban una ley potencial. Dave es más un pensador que un orador. «Yo no lo creo», dijo al final, y el «yo» se alargó como si durara media docena de sílabas, rezumando duda cada una de ellas. «En primer lugar», continuó, «los datos son horribles». Pero son tus datos, dije, sorprendido. «Lo sé, y no me gusta ir por ahí diciendo que mis datos son horribles. Son los mejores que tenemos, pero a uno le gustaría tener algo mucho mejor si pretende sacar conclusiones de esa magnitud». Los datos en cuestión registran medidas de niveles de extinción en setenta y nueve puntos durante la vida poscámbrica. Dave y su colega Jack Sepkoski han pasado años compilándolos, a veces juntos, a veces por separado. Es un trabajo que consume mucho tiempo, muy difícil, y la calidad del producto final refleja los problemas de la compilación, no la calidad de los investigadores. «Algún día dispondremos de datos lo bastante buenos como para hacer este tipo de cosas».
Pero supón, dije, que los datos fueran buenos ahora, y vieras una curva como la que Stu ha conseguido, algo cercano a la ley exponencial. ¿Sería válida su interpretación? «Me gano la vida con las simulaciones», empezó a modo de respuesta, «y sé lo delicado que es el proceso, lo plagado de trampas que está. A veces encuentras algo que refleja el mundo real de algún modo y piensas: “Eh, aquí me acerco a algo”. Es algo gestáltico y es seductor». Me habló de un nevado fin de semana de invierno que pasó en el Laboratorio de Biología Marina de Woods Hole, en Massachuchetts, muchos años atrás, con Steve Gould, Dan Simberloff (un ecólogo), Jack Sepkoski y otros, discutiendo con la esperanza de obtener alguna idea provechosa sobre los problemas de la biología evolutiva. No salía nada. Entonces, el domingo por la tarde, Dave sugirió que consideraran algunos de esos procesos como si fueran aleatorios. La sugerencia produjo una docena de artículos científicos en los años siguientes. Algunos buenos, otros muy malos.
«Steve, Dan y yo hicimos una simulación en la que asignamos iguales probabilidades de extinción a un biota modelo, luego buscamos pautas de extinción», me contó Dave. «Conseguimos pautas como las que ves en el registro fósil: grupos que crecen y disminuyen, algunos extinguiéndose al final, como en el mundo real». Era un momento en que cierto número de investigadores intentaba separarse del postulado de la biología evolutiva según el cual «todo está determinado por la inexorable dinámica de la selección natural». Fue una gran sorpresa que las pautas de las grandes extinciones pudieran producirse usando un enfoque puramente aleatorio. También estaba equivocado. «Nos equivocamos en la escala», explicó Dave. «Nuestros grupos eran demasiado pequeños, como señaló con razón Steve Stanley. Si se utilizan grupos mucho más grandes, no se obtiene en absoluto esa pauta. Así que, ya ves, me es familiar lo fácil que resulta equivocarse, sobre todo cuando algo parece tan irresistible».
¿Se está equivocando Stu?, pregunté. «En estadística hay algo que se llama el modelo del palo roto», dijo Dave. Un generador de números aleatorios «rompe» un palo de longitud 100 en veinticinco puntos y produce veintiséis palos más cortos. Los pedazos se miden, se cuentan los que miden una unidad, los que miden dos, etcétera y se dibuja un histograma. El resultado es una distribución asimétrica, desviada hacia el lado corto, como muchos fenómenos naturales, incluyendo la distribución de los tamaños de las ciudades estadounidenses, por ejemplo. «Algo que no hay que olvidar en el tema de las extinciones es que algunas especies son más propensas a desaparecer que otras, porque existen como pequeñas poblaciones aisladas», explicó Dave. «Este tipo de rareza estadística puede fácilmente sesgar los resultados». ¿Así que mostrarías tus reservas hacia cualquier cosa que se pareciera a una ley potencial? «Sí, porque es muy común, está en la naturaleza de la estadística. Puede decirte que un sistema está en equilibrio en el punto crítico, sea lo que sea lo que eso significa, pero puede que no. En cualquier caso, cuando Stu afirma que la curva que obtiene de mis datos se acerca a una ley potencial, sabe que hay otros muchos modelos matemáticos que podrían encajar perfectamente».
No cabía duda de que había muchas razones para mostrarse cauto al sacar la conclusión de que, usando sólo los datos de las extinciones, los ecosistemas globales están en equilibrio en el límite del caos. «Me has preguntado cómo interpretaría una distribución potencial válida para los tamaños de las extinciones», dijo Dave, volviendo a la pregunta original. «Bueno, ya sabes que sostengo que una fracción muy grande de las extinciones pudieron ser causadas por colisiones de asteroides. Y ya sabes que el tamaño de los asteroides puede describirse mediante una ley potencial: los grandes son escasos, los pequeños son comunes; es algo que se puede apreciar en la distribución del tamaño de los cráteres de la Luna. De manera que podría ser que una distribución potencial del tamaño y la frecuencia de la extinción sea un reflejo de una distribución potencial del tamaño y la frecuencia del impacto de asteroides. Podría ser, ¿verdad?».
«Sí, podría ser», admitió Stu cuando le hice la pregunta de Dave. ¿Cómo se sabe entonces si la distribución potencial de las extinciones está causada por la distribución de tamaño de los asteroides o si los ecosistemas globales están en equilibrio cerca del límite del caos? «Le pregunté a Per [Bak] qué sucede cuando se superpone una ley potencial sobre otra, a qué se parecería», dijo Stu. «Me respondió: “Seguirás viendo una ley exponencial”. Seguí preguntándole cómo, pero no llegué a ninguna parte. Creo que es bastante confuso». Stu también admitió que la curva de los datos de extinción podía describirse con otros modelos matemáticos, no sólo la ley potencial. «Pero, al menos, la curva es consistente con los ecosistemas globales que están cerca del límite del caos», dijo. «Sí, sé que eso es bastante débil. Pero, mira, si la curva no se pareciera en nada a una ley potencial, no tendríamos nada de qué hablar. En ella no aparecería el límite del caos. Tal como están las cosas, sigue siendo posible que los ecosistemas globales se coloquen en el límite del caos, como predicen nuestros modelos».
Por último, le pregunté a Stu por la conectividad. El modelo coevolutivo construye conexiones entre las especies del ecosistema. Es parte del sistema, la parte que se adapta al moverse el propio sistema hacia el límite del caos. La conectividad es necesaria si el sistema tiene que funcionar como conjunto, no sólo como suma de entidades independientes. Y la conectividad es necesaria si las perturbaciones tienen que caer en cascada por el sistema, produciendo avalanchas de especiación y extinción. En una breve incursión por la biología del problema, Dave había expresado sus dudas de que la conectividad en el mundo real fuera suficiente para propagar las consecuencias de perturbaciones mundiales y causar, así, extinciones en masa a escala global. ¿Tiene razón Dave al mostrarse escéptico?, pregunté. «Eso da que pensar», replicó Stu. «No tengo inconveniente en concebir una conectividad a escala continental, pero globalmente es más discutible. Necesitas conexiones a lo Gaia».
Mientras consideraba el problema, añadió: «Sí, para conseguir una extinción del 96 por ciento, como la de finales del Pérmico, hace falta mucha conectividad». Espera un momento, dije, ¿sabes que la extinción del Pérmico coincidió con la unión de los continentes, la formación de Pangea? «¿En serio? Vaya, fantástico».
Los continentes del planeta están en un movimiento constante y apenas perceptible, pasajeros de una fina corteza dividida en múltiples placas. Uno de los grandes descubrimientos de la ciencia del siglo XX, el movimiento continental como resultado de la tectónica de placas, arroja una nueva luz sobre la historia, una luz muy difícil de comprender para las mentes humanas, tan esclavas del presente. Inmersos en un deslizamiento global, los continentes a veces chocan y ocasionalmente se unen en un supercontinente, Pangea. La última vez que ocurrió fue a finales del Pérmico, hace 250 millones de años. «Eso significaría que todos los biotas estuvieron en contacto potencial entre sí, todos», dijo Stu.
Las colisiones y la unión de todos los continentes se han invocado, de hecho, como factores contribuyentes, cuando no causantes, de la extinción de finales del Pérmico. La argumentación es que si las masas terrestres se juntan para formar un continente gigante, se pierde alrededor de la mitad de la línea costera. (Basta hacer cuatro cuadrados de cartulina de cinco centímetros de lado y sumar el total de los perímetros; luego se unen formando un único cuadrado y se vuelve a medir el perímetro). La extinción a gran escala en el ámbito marino también es probable que se deba a este hecho. «Eso es cierto, por supuesto», dijo Stu. «Pero también proporciona el potencial de conectividad para las avalanchas coevolutivas a lo largo de toda la masa de tierra, para contribuir a la mayor extinción de la historia de la Tierra, ¿verdad?». Nos habíamos adentrado bastante en el terreno especulativo y cualquier huella de complejidad que pudiéramos vislumbrar tenía que ser considerada con un elevado grado de escepticismo.
No obstante, había visto huellas en otro sitio. Confusas y discutibles, cierto. Nada que permitiera justificar la declaración de que, en la explosión cámbrica y las extinciones en masa, la complejidad triunfa como fuerza dominante. Pero suficientes para alentar una mayor exploración de los patrones biológicos. Sabía que tenía que descender de las alturas de las grandes pautas y observar más de cerca los propios ecosistemas, tanto los reales como los que viven sólo en los ordenadores.