FIGURA 5.1. El dilema del prisionero.
¿De dónde viene todo el diseño?
—Perdone señor; ¿podría decirme cómo llegar a la sala de conciertos?
—¡Práctica, práctica y más práctica!
La Orquesta Sinfónica de Boston (OSB) es famosa por hacerles la vida imposible a los directores invitados hasta que demuestran su valía. Un joven director, a punto de debutar en la OSB y conocedor de la reputación de dicha orquesta, decidió ensayar un atajo para ganarse su respeto. Estaba previsto que dirigiera el estreno de una pieza contemporánea terriblemente discordante y, mientras repasaba la partitura, se le ocurrió una brillante estratagema. Encontró un crescendo hacia el comienzo de la pieza en el que la orquesta entera tocaba con toda su furia al menos doce notas discordantes y observó que el segundo oboe, una de las voces más suaves de la orquesta, debía tocar un si. Tomó esa parte de la partitura para el segundo oboe, e insertó cuidadosamente un signo de bemol: ahora el oboe tenía instrucciones de tocar un si bemol. En el primer ensayo, llevó briosamente a la orquesta hacia el crescendo previsto. «¡No!», gritó de repente, al tiempo que ordenaba parar a la orquesta. Luego, con la frente arrugada y mostrando una profunda concentración, dijo: «Alguien, veamos, sí, tiene que ser… el segundo oboe. Se suponía que debía tocar un si y usted ha tocado un si bemol». «Ni mucho menos —dijo el segundo oboe—, yo toqué un si. ¡Algún idiota había escrito un si bemol!».
Consideremos el fenómeno desde el punto de vista biológico. La Orquesta Sinfónica de Boston lleva existiendo más de un siglo, durante el cual su personal no ha dejado de cambiar, sus finanzas han sufrido altibajos, su repertorio ha ido creciendo y modificándose a medida que se han ido retirando piezas viejas y explorando otras nuevas. En muchos aspectos esta vieja institución es como un organismo vivo, con una personalidad marcada, una historia particular de crecimiento, salud y enfermedad, aprendizaje y olvido, viajes por todo el mundo y retornos a casa, sustitución de «células» viejas y cansadas por nuevos reclutamientos, adaptación de su comportamiento al nicho ecológico que le ha permitido prosperar. Esta perspectiva biológica resulta atractiva y útil, pero deja fuera los rasgos más importantes y asombrosos del fenómeno. Si los biólogos de otra galaxia descubrieran la Orquesta Sinfónica de Boston, lo que más debería impresionarles no son sus notables parecidos con los animales vías plantas, sino aquello que la diferencia de ellos. Un organismo está hecho de un gran equipo de células, pero ninguna célula puede sufrir ante la perspectiva de verse humillada. Ninguna célula puede aprender a tocar el oboe, o ser responsable de la elección del director invitado de este año entre una lista de jóvenes promesas. Ninguna célula puede comprender las implicaciones de la respuesta del oboísta y anticipar el efecto catastrófico que tendrá sobre la campaña del joven director para ganarse el respeto de la orquesta.
Lo notable de la Orquesta Sinfónica de Boston (y de la miríada de instituciones y prácticas humanas) es que, por un lado, puede llegar a ser una maravilla de diseño y organización, de autosuficiencia, mientras que, por otro lado, está compuesta por una multiplicidad de individuos autónomos, de diferentes nacionalidades, edades, géneros, temperamentos y aspiraciones. Los miembros de la orquesta son libres para ir y venir a su antojo, de modo que el consejo de dirección debe esforzarse por garantizar las condiciones laborales y pagar lo suficiente como para mantener motivados a los miembros de la orquesta. Fijémonos en la sección de los violines. Veinte individuos de talento, pero todos diferentes. Algunos son brillantes pero perezosos, mientras que otros son unos perfeccionistas obsesivos; uno está aburrido pero trabaja a conciencia, otro está embriagado por la música, mientras que otro está soñando despierto con la posibilidad de hacer el amor con la adorable celista de delante, pero todos ellos pasan sus arcos sobre las cuerdas perfectamente al unísono, una pauta impuesta con firmeza sobre un caleidoscopio de conciencias humanas distintas. Lo que hace posible esta acción concertada es un inmenso complejo de productos culturales profundamente compartidos por los músicos, la audiencia, el compositor, los conservatorios, los bancos, las autoridades municipales, los constructores de violines, las agencias de venta de entradas, y demás. Nada en el mundo animal puede acercarse a esta complejidad. Las mentes humanas están amuebladas —y acosadas— por miles de anticipaciones, evaluaciones, proyectos, planes, esperanzas, miedos y recuerdos que son totalmente inaccesibles a las mentes de nuestros parientes más próximos, los primates superiores. Este mundo de ideales y creaciones humanas proporciona a los seres humanos individuales unas capacidades y unas tendencias que son marcadamente distintas a las de cualquier otro ser vivo del planeta.
La libertad del pájaro para volar allí donde se le antoje es sin duda un tipo de libertad, una mejora incuestionable respecto a la libertad de la medusa de flotar por donde flota, pero un triste primo lejano de la libertad humana. Compárese el canto de los pájaros con el lenguaje humano. Ambos son unos productos magníficos de la selección natural, y ninguno de los dos es milagroso, pero el lenguaje humano revoluciona la vida, abre el mundo biológico a dimensiones completamente inaccesibles para los pájaros. La libertad humana, producto en parte de la revolución del lenguaje y la cultura, es tan diferente de la libertad del pájaro como pueda serlo el lenguaje del canto de los pájaros. Pero para comprender el fenómeno más rico, es necesario comprender primero sus componentes y predecesores más modestos. Lo que debemos hacer para comprender la libertad humana es seguir la «extraña inversión del razonamiento» de Darwin y remontarnos a la época del origen de la vida, cuando no había libertad, ni inteligencia, ni elección, sino sólo una protolibertad, una protoelección, una protointeligencia. Disponemos ya de un cuadro general de lo que ocurrió: células simples dieron origen con el tiempo a células más complejas, las cuales dieron origen a su vez a organismos multicelulares, los cuales dieron origen a su vez al complejo mundo macroscópico en el que vivimos y actuamos. Ahora debemos remontarnos y examinar algunos de los detalles más relevantes de esta procesión.
Supongamos que lo que queremos es estar vivos en el planeta Tierra. ¿Qué necesitamos? Si comenzamos a nivel molecular, no sólo necesitamos ADN, sino todas las herramientas —proteínas— moleculares para llevar a cabo los muchos pasos que requiere la replicación del ADN. Necesitamos una proteína para iniciar el proceso, otra para desenrollar la hélice, otra para fijar la cadena separada de ADN, etc., relajar el superenrollamiento, segmentar y agrupar los cromosomas, y así sucesivamente. Ninguno de estos pasos es opcional; todos son necesarios. Si falta alguna de estas proteínas, adiós. Esos mismos materiales de construcción tuvieron que ser diseñados a lo largo del tiempo. El equipo completo, que compartimos con toda la vida actual del planeta, se fue reuniendo y refinando a lo largo de varios miles de millones de años, y sustituye a equipos más sencillos para nuestros ancestros aún más sencillos. Dependemos de nuestro equipo, y ellos dependían del suyo, pero tenemos más posibilidades que ellos gracias a las mejoras introducidas en nuestro equipo, mejoras que han sido posibles gracias a formas de integración más elevadas, las cuales a su vez han sido posibles gracias a formas aún más astutas de colisionar con las demás cosas del mundo y explotar los resultados de dichas colisiones. Cuando comenzó la vida, había sólo una forma de estar vivo. Se trataba de hacer A o morir. Ahora hay opciones: hacer A o B o C o D o… morir.
Para vivir se necesita energía. ¿Procedía del sol la primera energía explotada para la vida o de fuentes termales situadas en las profundidades de la Tierra? Esta cuestión sigue abierta en la actualidad y ha dado pie a una cantidad abrumadora de hipótesis sobre el origen de la vida que compiten por encontrar una confirmación. Fuera cual fuera su comienzo, la vida —o, en cualquier caso, la mayor parte de la vida— terminó por depender de la energía solar. Para mantenerse vivo y reproducirse era preciso estar cerca del mar o flotar en su superficie, tomando el sol. Una gran innovación se produjo cuando algunos de estos bañistas mutaron y, de este modo, «descubrieron» que, en lugar de hacerlo todo ellos mismos, podían obtener mejores resultados si invadían y desmembraban a algunos de sus vecinos para usarlos como un práctico almacén de piezas de repuesto ya construidas. Las irrupciones son lo que hace la vida interesante. Invasores e invadidos inauguraron una carrera de armamentos que llevó a nuevas variedades de ambos. Pronto —al cabo de mil millones de años aproximadamente— había muchas «maneras de ganarse la vida» (por usar la expresión de Richard Dawkins), pero esas muchas no serán nunca más que una desvaneciente cadena de actualizaciones en medio del Vasto espacio de posibilidades lógicas. Casi todas las combinaciones del material de construcción son una forma de no estar vivo.
Una de las innovaciones más importantes dentro de esta carrera armamentística de diseño competitivo fue el accidente conocido como la revolución eucariótica, que tuvo lugar hace varios miles de millones de años. Los primeros seres vivos, las células relativamente sencillas conocidas como procariotas, tuvieron todo el planeta para ellas durante unos tres mil millones de años, hasta que una de ellas fue invadida por una vecina, y el equipo resultante de dos fue más apto que sus primos no infectados, de modo que prosperaron y se multiplicaron, y transmitieron su capacidad de trabajar en equipo a sus descendientes. Era un primer ejemplo de un cierto tipo de cooperación: la simbiosis, un caso en el cual se produce una colisión entre X e Y, pero en lugar de que X destruya a Y, o al revés, o incluso peor, que ambos resulten destruidos —el resultado habitual de las colisiones en este duro mundo—, X e Y unen sus fuerzas y crean Z, una nueva cosa más grande y versátil, con más opciones de salir adelante. Por supuesto, es posible que esto ocurriera muchas veces en el mundo procariota, pero desde la primera vez que ocurrió, la vida en el planeta cambió para siempre. Estas nuevas supercélulas, las eucariotas, vivían junto a sus primas procariotas, pero eran mucho más complejas, versátiles y competentes gracias a sus polizones. Se trataba de una cooperación involuntaria, sin duda. Los equipos eucariotas no tenían la menor noticia del trabajo en equipo que estaban realizando. No teman —ni necesitaban— ninguna noción del arbitrario origen de su ventaja competitiva. Los primeros seres vivos eucariotas no eran multicelulares, pero abrieron un espacio de diseño que hacía posibles los organismos multicelulares, puesto que tenían las suficientes piezas de recambio como para convertirse en diferentes clases de especialistas. (Todavía estamos muy lejos de los violinistas y los oboístas, y del trabajo en equipo de la OSB, pero vamos por el buen camino).
La revolución eucariótica nos lleva a fijarnos en el hecho de que incluso en la evolución biológica, que Darwin llamó adecuadamente «descendencia con modificación», hay mucho espacio para la transmisión horizontal del diseño. Los anfitriones procariotas que fueron los primeros «infectados» por sus simbióticos visitantes recibieron un inmenso regalo para mejorar su competencia, el cual había sido diseñado en otro lugar. Es decir, no recibieron toda su competencia por línea vertical de sus ancestros, a través de sus padres, sus abuelos y sus demás antepasados. En otras palabras, no recibieron toda su competencia de sus genes. Lo que sí hicieron, sin embargo, fue transmitir este regalo a toda su descendencia posterior a través de sus genes, ya que los genes de los invasores terminaron por compartir el destino de los genes del núcleo de sus anfitriones, y viajaron unos al lado de los otros hacia la siguiente generación, que fue infectada al nacer, podría decirse, con su propio complemento simbiótico. Hay un rastro claro de este doble camino que sigue siendo aún muy visible hoy en día en todas las criaturas multicelulares, incluidos nosotros. Las mitocondrias, los minúsculos organillos que transforman energía dentro de nuestras células, son los descendientes de aquellos invasores simbióticos y tienen sus propios genomas, su propio ADN. Nuestro ADN mitocondrial, que recibimos únicamente de nuestra madre, existe en cada una de nuestras células junto con nuestro ADN nuclear (nuestro genoma). (La reproducción sexual llegó mucho más tarde; el esperma del padre no aporta nada de sus mitocondrias en el proceso de fertilización).
La transmisión horizontal del diseño, de información que puede ser aplicada a fines útiles, es la clave de la cultura humana, y sin duda el secreto de nuestro éxito como especie. Cada uno de nosotros es el beneficiario del trabajo de diseño realizado por incontables individuos que no son ascendientes nuestros. Cada uno de nosotros no se ve obligado a «reinventar la rueda», o el cálculo, los relojes o la forma soneto. A veces se pretende, erróneamente, que esta transmisión cultural entre individuos que no guardan lazos genéticos demuestra que la cultura humana no puede interpretarse como un fenómeno evolutivo gobernado por los principios de la teoría neodarwiniana. En realidad, tal como acabamos de ver, la transmisión horizontal de elementos útiles de diseño entre individuos no emparentados está reconocida como un elemento importante en la evolución de las formas más tempranas de vida (unicelular) del que existe una lista cada vez más amplia dé ejemplos demostrados y constituye, por lo tanto, una pieza central, no un motivo de vergüenza, para la biología evolutiva contemporánea.
La revolución eucariótica no tuvo lugar de un día para otro; antes de que pudiera asentarse fue necesario que la evolución descubriera laboriosamente las soluciones a muchos problemas. Ya en el capítulo 2 encontramos los parásitos transposons, genes renegados cuyos efectos había que frenar. El proceso que resolvió dichos conflictos intragenómicos ilustra varios aspectos importantes de la teoría darwiniana: la I+D es cara, hay que «pagar» por cada diseño, y la evolución recicla permanentemente diseños anteriores (va pagados y copiados) para nuevos fines. Los genes de una simple célula procariota pueden expresarse con ayuda de un equipo relativamente sencillo de lectura de genes. Es decir, no se requiere una tecnología muy desarrollada para seguir la receta de unos genes procariotas y construir un nuevo procariota. Las más sofisticadas células eucariotas, en cambio, y no digamos los organismos multicelulares compuestos de dichos bloques de construcción más complejos, necesitan un sistema de pasos intermedios, controles y equilibrios abrumadoramente elaborados para que los genes puedan activarse y desactivarse en los momentos adecuados gracias al efecto indirecto de otros productos genéticos, y así sucesivamente. Durante algún tiempo, los biólogos se han enfrentado al clásico dilema del huevo y la gallina: ¿cómo pudo evolucionar esta elaborada maquinaria de regulación de genes? La vida multicelular no podía ni siquiera iniciar su evolución antes de que estuviera lista la mayor parte de esta maquinaria, pero aparentemente no es necesaria para la forma más simple de vida de las procariotas. ¿Qué fue lo que pagó toda esta I+D? La respuesta que comienza a ganar fuerza es que fue costeada por una guerra civil de unos mil millones de años al comienzo de la vida procariota. Era una carrera de armamentos dentro del propio genoma, en la que los genes cívicos luchaban contra los transposons (unos gorrones que se copiaban una y otra vez en el genoma sin aportar ningún beneficio al conjunto del organismo). Esto provocó la aparición de gran cantidad de medidas y contramedidas, tales como mecanismos de silenciación y mecanismos antiaislamiento. (Cada vez conocemos mejor los detalles de dichos mecanismos, así como los detalles de los mecanismos que permitieron las unificaciones simbióticas de genomas en la revolución eucariótica, y son sin duda fascinantes, pero van más allá del alcance de este libro). Igual que en las carreras armamentísticas actuales, el resultado fue un empate muy caro, pero los frutos de toda aquella I+D estaban disponibles para abrir nuevos caminos: la maquinaria de alta tecnología necesaria para crear formas de vida multicelulares (McDonald, 1998). Parece, pues, que nosotros somos una especie de «dividendo de la paz», igual que los ordenadores, el Teflon, el GPS y tantos otros frutos tecnológicos de la carrera armamentística dirigida por el complejo militar industrial con los dólares de nuestros impuestos.
Pero ¿cómo son estas carreras de armamentos? ¿Qué factores gobiernan o limitan los ataques y contraataques de los diferentes «bandos» en estas competiciones? Cada vez que surge en la naturaleza algo parecido a la cooperación es preciso encontrar una explicación. (Es posible que tenga su origen en un feliz accidente, pero no puede ser que se mantenga por un feliz accidente. Eso sería demasiado bueno para ser cierto). Aquí es donde necesitamos la perspectiva de la teoría de juegos, y de su ejemplo clásico, el dilema del prisionero. Se trata de un sencillo «juego» para dos personas que lanza algunas pistas, unas evidentes y otras sorprendentes, sobre diferentes aspectos de nuestro mundo. El escenario básico es el siguiente. Ha sido usted encarcelado junto a otra persona en espera de juicio (digamos que bajo una falsa acusación) y el fiscal les ofrece a ambos, por separado, el mismo acuerdo: si callan los dos, sin confesar ni implicar al otro, les caerá a ambos una sentencia corta (las pruebas del Estado no son demasiado concluyentes); si usted confiesa e implica al otro y este calla, queda usted en libertad, y a el le cae cadena perpetua; si ambos confiesan e implican al otro, ambos obtienen sentencias intermedias. Por supuesto, si usted calla y la otra persona confiesa, él sale libre y a usted le cae cadena perpetua.
Si ambos se niegan a colaborar, desafiando al fiscal, el resultado sería mucho mejor para los dos que si ambos confesaran, de modo que ¿por qué no hacerse la promesa de callar? (En la jerga habitual del dilema del prisionero, la opción de callar se llama cooperar —con el otro prisionero, por supuesto, no con el fiscal—.) Podrían prometérselo, pero ambos tendrían la tentación —cedieran o no a ella— de traicionar, puesto que en tal caso cada uno saldría libre, dejando al otro primo con todo el lío. Como se trata de un juego simétrico, la otra persona estará igual de tentada, por supuesto, de hacerle quedar a usted como un tonto con su traición. ¿Está usted dispuesto a arriesgar toda una vida en la cárcel a que el otro mantendrá su promesa? Tal vez sea más seguro traicionar, ¿verdad? De ese modo evita usted en todo caso el peor de los resultados, y tal vez incluso salga libre. Por supuesto, el otro tipo también llegará a esta conclusión, si es una idea tan buena, de modo que probablemente también optará por lo seguro y le traicionará, en cuyo caso debe usted traicionarlo para evitar la peor de las calamidades —a menos que sea un santurrón dispuesto a pasar el resto de la vida en la cárcel para salvar a un traidor—, por lo que es probable que ambos terminen por traicionarse y recibir sentencias intermedias. ¡Qué bonito sería poder superar esta línea de razonamiento y cooperar!
Lo importante es la estructura lógica del juego, no su escenario concreto, que no es sino un estímulo para la imaginación. Podemos sustituir las sentencias penales por resultados positivos (oportunidades de ganar diferentes cantidades de dinero o, digamos, de descendientes) mientras las retribuciones sigan siendo simétricas y estén ordenadas de tal modo que una traición solitaria sea más beneficiosa que la cooperación mutua, la cual dé mejores resultados que la traición mutua, la cual, a su vez, dé mejores resultados que la posición del primo, que se produce cuando el otro es el único traidor. (En entornos formales se establece otra condición: la media de las retribuciones del primo y la traición mutua no debe ser mayor que la retribución de la cooperación mutua). Siempre que se cumpla esta estructura en el mundo, nos encontramos ante el dilema del prisionero.
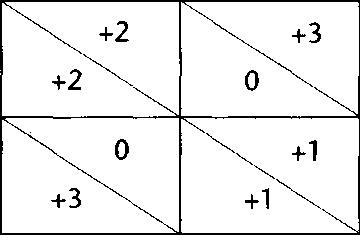
Se han emprendido estudios teóricos a partir de juegos en los campos más diversos, como la filosofía, la psicología, la economía y la biología. En la teoría de juegos evolutiva, las retribuciones se miden en términos de descendientes y el objetivo de los modelos es examinar las condiciones bajo las cuales los diseños «cooperativos» pueden mantenerse y superar en resultados a las instancias egoístas y traidoras que habitualmente se llevan la mejor parte. ¿Por qué es la traición la estrategia ganadora por defecto? Examinemos la matriz de retribuciones de la figura 5.1. Haga lo que haga el jugador Y, si el jugador X traiciona, obtendrá mejores resultados que si coopera. Se dice que la traición domina como estrategia en la situación básica. Se puede derivar matemáticamente el efecto que ello tendrá sobre los descendientes del jugador X como proporción de la generación siguiente dentro de una determinada población, y demostrarlo en simulaciones que enfrentan a agentes traidores simples de diferentes tipos con agentes cooperadores simples de diferentes tipos. Ambos interactúan en función de su tipo —los traidores siempre traicionan y los cooperadores siempre cooperan— y los resultados (en términos de número de descendientes) se van contando y acumulando a lo largo de muchas generaciones. En ausencia de circunstancias especiales que lo impidan, los traidores pronto desbordan a los cooperadores, por desgracia. Esta tendencia inevitable es el viento dominante contra el que toda la evolución de la cooperación debe luchar. La más influyente de las muchas aplicaciones del pensamiento teórico sobre juegos a la teoría evolucionista es el concepto de estrategia evolutivamente estable, o EEE, de John Maynard Smith, una estrategia que tal vez no sea la mejor imaginable pero que resulta insuperable por cualquier otra estrategia alternativa bajo las circunstancias dadas. Un mundo sucio donde todos traicionan siempre es una EEE en la mayoría de las circunstancias imaginables, puesto que los cooperadores pioneros que van a parar en medio de una población de este tipo son traicionados hasta la muerte en poco tiempo. Existen condiciones, sin embargo, bajo las cuales se dan otros resultados más alentadores, y esas huidas del sombrío caso general son los peldaños de la escalera que lleva hasta nosotros.
FIGURA 5.1. El dilema del prisionero.
No hay duda de que los análisis de la teoría de juegos son útiles para la teoría evolucionista. ¿Por qué, por ejemplo, son tan altos los árboles en el bosque? Por lo misma razón por la que grandes despliegues de carteles chillones compiten por nuestra atención en los centros comerciales de todo el país. Cada árbol piensa sólo en sí mismo y trata de conseguir tanto sol como sea posible. Sería mucho mejor que esas secuoyas se pusieran de acuerdo en establecer unas cuantas restricciones zonales razonables y dejaran de competir entre ellas por la luz, lo que les permitiría ahorrarse la molestia de construir esos ridículos y caros troncos, no ir más allá de ser unos matorrales bajos y ahorrativos, ¡y conseguir el mismo sol que de la otra manera! Pero no pueden ponerse de acuerdo. Bajo esas circunstancias, es inevitable que la traición produzca siempre mejores resultados que cualquier acuerdo cooperativo, de modo que si no fuera porque el suministro de luz solar es esencialmente inagotable, los árboles se verían atrapados en la «tragedia de los comunes» (Hardin, 1968). La tragedia de los comunes se produce cuando hay un recurso finito «público» o compartido que los individuos se sentirán tentados de explotar de manera egoísta más allá de la parte que les corresponde (como por ejemplo los peces comestibles). A menos que se llegue a acuerdos específicos y coercibles, el resultado tenderá a ser la destrucción del recurso. Fue el desarrollo evolutivo de controles y equilibrios coercibles lo que permitió que los genes cooperadores resistieran frente a los transposones traidores, una de las primeras innovaciones «tecnológicas» para superar el mundo aburrido y simple del egoísmo universal, la traición universal.
El paso a la multicelularidad vino auspiciado por otra innovación en el terreno de la cooperación: resolver el problema de la solidaridad de grupo a nivel celular. Tal como señalé al comienzo del capítulo 1, cada uno de nosotros está compuesto de billones de células robóticas, cada una de las cuales dispone de un sistema completo de genes y una impresionante maquinaria interna de soporte vital. ¿Por qué se someten dichas células de manera tan altruista para el bien del equipo en conjunto? Por supuesto, lo que pasa es que han llegado a ser tremendamente dependientes las unas de las otras, y no pueden sobrevivir mucho tiempo por sí mismas fuera del entorno particular donde acostumbran a vivir; pero ¿cómo llegaron a ser así[3]? Una de las virtudes de adoptar el «punto de vista del gen» en los estudios evolutivos es que convierte esta cuestión en un problema fundamental. La solidaridad de grupo entre las células es omnipresente en la naturaleza; al fin y al cabo, cualquier ser vivo que podamos ver a ojo descubierto está formado por células serviles y devotas. Se trata, pues, de algo «natural», pero no por ello deja de ser una proeza del diseño de proporciones sumativas, nada que los biólogos puedan dar por supuesto. Las lecciones que debemos aprender de todo ello son engañosas, sin embargo, ya que las células que nos componen pertenecen a dos categorías muy distintas.
Todas las células que componen un yo multicelular comparten un mismo ancestro; pertenecen a un único linaje celular, son hijas y nietas de un óvulo y un esperma que se unieron para formar un zigoto. Esas son las células hospedadoras. Las demás células, los simbiontes, son como las otras —son también eucariotas y procariotas—, pero cabe considerarlas forasteras porque descienden de linajes distintos. (Así pues, tenemos una simbiosis de segunda generación; la simbiosis creó nuestras células eucariotas, las cuales a su vez han hecho de anfitrionas para un sinfín de nuevos invitados).
¿Qué es lo que conlleva la distinción entre anfitriones e invitados? La respuesta aquí, que encontrará eco en los niveles más elevados de la vida social, es que aunque el pedigrí es a menudo una buena manera de predecir la competencia futura, al final lo que cuenta es la competencia futura, con independencia del pedigrí. Por ejemplo, nuestro sistema inmunológico está compuesto por células que son ahora mismo miembros de pleno derecho del equipo anfitrión, pero iniciaron su carrera con nuestros antepasados como un ejército invasor, que fue gradualmente incorporado y convertido en una tropa de guardias mercenarios, cuya identidad genética se mezcló con los linajes más antiguos a los que unieron sus fuerzas, en otro ejemplo de transmisión horizontal del diseño. La clave para comprender las pautas que siguen estas transformaciones es tratar a todas estas células robóticas como agentes individuales minúsculos, como sistemas intencionales, cada uno de los cuales posee un principio de capacidad de elección «racional». Adoptar la perspectiva intencional, pasar de la perspectiva física de los átomos componentes a la perspectiva del diseño de las simples máquinas primero, y luego a la perspectiva intencional de la agencia simple, es una táctica que da grandes resultados pero que debe utilizarse con precaución. Resulta demasiado fácil perder de vista el hecho de que hay momentos en las carreras de todos estos agentes, semiagentes y pseudosemiagentes en los que surgen —y luego pasan— oportunidades para «decidir».
Las células que me componen comparten un mismo destino, aunque algunas lo hacen en un sentido más fuerte que otras. El ADN de las células de mi dedo y mis células sanguíneas están en un callejón sin salida genético; dichas células pertenecen a la línea somático (el cuerpo), no a la línea germinal (las células sexuales). Según la memorable expresión de François Jacob, el sueño de toda célula es convertirse en dos células, pero las células de mi línea somática están condenadas a morir «sin hijos» (aparte de la posibilidad de aportar ocasionalmente un sustituto para sus vecinas muertas en acción, y de posibles avances sin precedentes en las técnicas de clonación). Como este callejón sin salida se determinó hace ya bastante tiempo, ya no hay ninguna presión, ninguna oportunidad normal, ningún «punto de elección» donde sus trayectorias intencionales —o las trayectorias de su limitada progenie— puedan alterarse. Son lo que podría llamarse un sistema balístico intencional, cuyos objetivos y propósitos superiores han sido fijados de una vez por todas, sin ninguna posibilidad de reorientación o variación. Son esclavos completamente sometidos al summum bonum del cuerpo del que forman parte. Cabe la posibilidad de que sean explotados o engañados por visitantes externos, pero bajo circunstancias normales no pueden rebelarse por sí mismos. Igual que a las Esposas Stepford, se les ha inculcado un único summum bonum, que no es: «Lucha por ser el número uno». Al contrario, son jugadores de equipo por naturaleza.
Cómo deben contribuir a este summum bonum es algo que viene implícito en su diseño, y en este sentido son fundamentalmente distintos de otras células que están «en el mismo barco»: mis visitantes simbiontes.
Los mutualistas benignos, los comensales neutrales y los parásitos nocivos que comparten el vehículo que componen entre todos —a saber, yo— tienen todos un summum bonum implícito en su diseño y su fin es promover sus propias cepas, no la mía. Por fortuna, se dan condiciones bajo las cuales puede mantenerse una entente cordiale, ya que, después de todo, estamos todos en el mismo barco vías condiciones bajo las cuales pueden obtener mejores resultados sin colaborar son limitadas. Pero sí tienen posibilidad de «elección». Es una cuestión abierta para ellas, a diferencia de lo que ocurre, normalmente, con las células hospedadoras.
¿Por qué? ¿Qué es lo que permite —o requiere— que las células hospedadoras sean tan serviles y al mismo tiempo den libertad a las células visitantes para rebelarse cuando surja la oportunidad de hacerlo? Por supuesto, ninguna célula es un agente racional capaz de pensar y sentir. Y ninguna posee capacidades cognitivas significativamente superiores a las demás. No es esa la base de la teoría de juegos de la evolución. Las secuoyas tampoco son especialmente inteligentes, pero se encuentran en unas circunstancias competitivas que les obligan a traicionar, y a caer en lo que, desde su punto de vista (!), no es sino un desperdicio inútil. El acuerdo de cooperación mutua por el que todas podrían evitarse construir troncos elevados en el vano intento de conseguir más luz de la que les corresponde es evolutivamente incoercible.
La condición que genera la posibilidad de una elección es el «voto» inconsciente de la reproducción diferencial. Es la oportunidad de que se produzca una reproducción diferencial lo que permite que las cepas de nuestros visitantes «cambien de idea» o «reconsideren», al «explorar» estrategias alternativas, las opciones que tienen. Mis células hospedadoras, en cambio, han sido diseñadas de una vez para siempre por un único voto en el momento de la formación de mi zigoto. Si, gracias a una mutación, se les ocurren estrategias dominantes o egoístas, no prosperarán (en relación con sus contemporáneas), puesto que hay escasas oportunidades para la reproducción diferencial. (El cáncer puede verse como una rebelión egoísta —y destructora del vehículo— promovida por una revisión de las circunstancias normales a partir de la cual se hace posible la reproducción diferencial).
Brian Skyrms ha señalado (1994a, 1994b) un sugestivo paralelismo entre esta estrategia multicelular (otro fruto benigno de la guerra civil que creó toda la maquinaria para la lectura de genes) y la monumental Teoría de la justicia (1971) de John Rawls. Una cooperación normal en el marco del destino estrechamente compartido de las células de la línea somática tiene como precondición una situación análoga a la que se da en la «posición original», el experimento mental de Rawls acerca de cómo diseñarían el Estado justo ideal unos agentes racionales si tuvieran que elegir desde detrás de lo que él llama un «velo de ignorancia». Skyrms lo llama, no sin razón, el «velo darwiniano de ignorancia». Nuestras células sexuales (esperma u óvulo) se forman por un proceso distinto de la división celular normal, o mitosis. Nuestras células sexuales surgen de un proceso diferente llamado meiosis, un proceso que construye aleatoriamente medio genoma (candidato a unir sus fuerzas con la otra mitad aportada por nuestro compañero o compañera) a base de elegir primero un fragmento de la «columna A» (los genes que recibimos de nuestra madre) y luego un fragmento de la «columna B» (los genes que recibimos de nuestro padre) hasta que la célula sexual tiene instalado un conjunto completo de genes —aunque sólo una copia de cada uno—, listo para probar suerte en la gran lotería del apareamiento. Pero ¿cuáles de las hijas de nuestro zigoto original están destinadas a la meiosis y cuáles a la mitosis? Esto también es una lotería.
¿Estamos hablando de una lotería aleatoria o pseudoaleatoria? Por lo que sabemos, viene a ser lo mismo que lanzar una moneda al aire, es decir, el resultado viene deterruinado por una coincidencia carente de pautas entre inescrutables influencias venidas de quién sabe dónde, y es por lo tanto predecible, en principio, por el demonio infinito de Laplace, pero no por las muy sensibles y variadas fuerzas selectivas que constituyen los ciegos pero efectivos tanteos del Relojero Ciego. Gracias a este mecanismo, los distintos genes paternos y maternos (en cada uno de nosotros) no pueden «conocer su destino» por adelantado en circunstancias ordinarias. La cuestión de si van a tener progenie en la línea germinal y tal vez inundar el futuro con descendientes suyos, o de si serán relegados a las aguas estériles de la esclavitud de la línea somática para el bien del cuerpo político o la corporación (nótese la etimología), es algo desconocido e imposible de conocer, de modo que los genes no tienen nada que ganar con una competición egoísta. Ese es, en todo caso, el arreglo más común. Hay ocasiones especiales en las que el Velo Darwiniano de la Ignorancia se levanta por un momento: los casos de «deriva meiótica» o de «impresión genómica» (Haig y Grafen, 1991; Haig, 1992, 2002; para una discusión véase La peligrosa idea de Darwin [Dennett, 1995, capítulo 9]), en los cuales las circunstancias sí permiten que tenga lugar una competición «egoísta» entre los genes —y eso es exactamente lo que ocurre, y lleva a una escalada de la carrera armamentista—. Pero en la mayoría de las circunstancias, el «tiempo del egoísmo», en el caso de los genes, está estrictamente limitado, y en cuanto el dado —o la papeleta— ha sido lanzado, los genes están fuera de la partida hasta la próxima elección. Tal vez el primero en señalar el paralelismo fue E. G. Leigh:
Es como si estuviéramos ante un parlamento de genes: cada uno actúa según su propio interés, pero si sus actos perjudican a los demás, estos se combinarán para frenarlo. Las reglas de transmisión de la meiosis han evolucionado hasta convertirse en unas reglas de fairplay cada vez más inviolables, una constitución diseñada para proteger al parlamento frente a los actos nocivos de uno solo o unos pocos. Sin embargo, en aquellos casos en los que la relación con el elemento distorsionador es tan estrecha que «seguir sus pasos» produce más beneficios que perjuicios, la selección tiende a potenciar la distorsión. Así, es necesario que una especie tenga muchos cromosomas para que, ante la aparición de un elemento distorsionador, la selección promueva su supresión en la mayor parte de los casos. Del mismo modo que un parlamento demasiado pequeño puede verse pervertido por las intrigas de irnos pocos, una especie que sólo tuviera un cromosoma con vínculos muy estrechos sería una presa fácil para los elementos distorsionadores (Leigh, 1971, pág. 249).
¡Que alguien trate de describir estas pautas profundas de la naturaleza sin usar la perspectiva intencional! Las lentísimas pautas que resultan predictivas al nivel de los genes recuerdan notablemente las pautas que se revelarán predictivas en los niveles psicológico y social, y en realidad son anticipos de ellas: las oportunidades, la ignorancia y el discernimiento, la búsqueda de las mejores maniobras frente a la competencia, la evitación y la represalia, la elección y el riesgo. Hay razones que explican las medidas y contramedidas de la I+D evolutiva, aunque nada o nadie las haya valorado explícitamente. Son lo que llamo rabones virtuales y precedieron en miles de millones de años a nuestras razones articuladas y ponderadas. Una de ellas es el principio fundamental de la evitación del daño, que rige en ambos dominios: cuando no tenemos información alguna acerca de cuál va a ser nuestro destino, la libertad de elección es perfectamente inútil.
Y todavía hay otra forma de negarles una determinada oportunidad a las personas: mantenerlas en la ignorancia de la misma. Podríamos llamar a estas oportunidades no reconocidas ni imaginadas oportunidades en bruto. Si paso caminando junto a una fila de contenedores de basura, y resulta que uno de ellos contiene un bolso lleno de diamantes, me pierdo una oportunidad en bruto de hacerme rico […]. Las oportunidades en bruto son muy abundan tes, pero no nos basta con ellas; cuando decimos que queremos oportunidades o posibilidades de prosperar, no queremos simplemente oportunidades en bruto. Queremos detectar nuestras oportunidades o ser informados de ellas a tiempo para que podamos actuar (Dennett, 1984, págs. 116117).
Skyrms demuestra que cuando los elementos individuales de un grupo —sea de organismos completos o de partes de ellos— están estrechamente relacionados (son clones o casi clones) o son de otro modo aptos para el reconocimiento o el «apareamiento» por semejanza, el modelo correcto para analizar la situación deja de ser el simple dilema del prisionero, en el que domina siempre la estrategia de la traición. Es por eso por lo que nuestras células somáticas no se traicionan; son clones. Esta es una de las condiciones para que los grupos —como el grupo de mis células «anfitrionas»— posean la armonía y la coordinación necesarias para comportarse de manera estable como un «organismo» o un «individuo». Pero antes de dar tres hurras y aceptarlo como nuestro modelo preferido para construir una sociedad justa, deberíamos detenernos a considerar que hay otra forma de ver a esos ciudadanos modelos, las células y los órganos de la línea somática: su modalidad específica de altruismo es la obediencia acrítica de los fanáticos, caracterizada por una lealtad al grupo fuertemente xenófoba que difícilmente podemos considerar un ideal para la emulación humana.
Nosotros, a diferencia de las células que nos componen, no nos encontramos en trayectorias balísticas; somos misiles guiados, capaces de alterar nuestra trayectoria en cualquier punto, abandonar objetivos, cambiar de lealtades, formarnos propósitos y luego traicionarlos, y demás. Para nosotros, cada momento es momento de decidir. Por este motivo, nos encontramos constantemente con la clase de oportunidades y dilemas sociales para los que la teoría de juegos propone un campo y unas reglas de juego, pero todavía no las soluciones. La vida es más complicada para la gente que vive en sociedad que para las células que los componen, y todavía nos queda mucha I+D que realizar —¡práctica, práctica y más práctica!— antes de llegar a la sala de conciertos.
Sin embargo, resulta alentador ver que los problemas a los que nos enfrentamos tienen precedentes que pudieron ser superados por ensayo y error. De otro modo no estaríamos aquí. El proceso de ensayo y error —incluso un ensayo y error inconsciente, capaz de conservar los progresos parciales— es poderoso. Ha sido capaz de crear novedades genuinas en el mundo; ha resuelto problemas de gran calado y superado obstáculos que harían retroceder a cualquiera. El método de ensayo y error funciona, lo que significa que probar funciona: al menos una de estas pruebas tiene un historial de éxitos más que probado. Tal vez nuestros intentos no parezcan tan torpes —a pesar del determinismo— a la vista del éxito que tuvieron sus antecesores. Las propias células que nos componen son los descendientes directos de células que una vez tuvieron que resolver un gran problema de cooperación, y lo lograron.
Después de esta siniestra disquisición sobre células y genes, combinada con otra disquisición sobre violinistas y oboístas, tal vez comience a ser hora de traer la paz a las conciencias desterrando de una vez para siempre el «espectro» del «determinismo genético». Según Stephen Jay Gould, los deterministas genéticos creen lo siguiente:
Si estamos programados para ser lo que somos, entonces dichos caracteres son ineluctables. Como máximo podemos canalizarlos de un modo u otro, pero no podemos cambiarlos ni por nuestra voluntad, ni por nuestra educación, ni por nuestra cultura (Gould, 1978, pág. 238).
Si esto es el determinismo genético, entonces podemos respirar todos aliviados: no hay deterministas genéticos. Nunca he encontrado a nadie que pretenda que ni la voluntad, ni la educación, ni la cultura no puedan cambiar muchos, si no todos, de nuestros rasgos genéticamente heredados. Mi tendencia genética a la miopía se ve compensada por las gafas que llevo (pero es preciso que quiera llevarlas); y muchas personas de las que de otro modo padecerían una u otra enfermedad de origen genético pueden ver sus síntomas pospuestos indefinidamente con sólo recibir la debida educación respecto a la importancia de una determinada dieta o gracias al don cultural de la prescripción de una u otra medicina. Si tenemos el gen causante de la enfermedad de la fenilcetonuria, todo lo que debemos hacer para evitar sus indeseables efectos es dejar de comer alimentos que contengan fenilalanina. Tal como hemos visto, lo inevitable no depende de si reina o no el determinismo, sino de si se pueden o no tomar medidas, basadas en información que podamos obtener a tiempo, para evitar el daño previsto. Para que una decisión tenga sentido es preciso que se cumplan dos requisitos: información y posibilidad de actuar en relación con esta información. Sin lo uno, lo otro es inútil o aún peor. En su excelente manual de genética contemporánea, Matt Ridley (1999) pone de relieve esta idea con el dramático ejemplo de la enfermedad de Huntington, que es «puro fatalismo, sin mezcla de variabilidad ambiental. Ni una buena vida, ni un buen tratamiento médico, ni una comida sana, ni una familia entregada, ni una gran fortuna pueden nada contra ella» (pág. 56). Se trata de un caso muy distinto de las muchas predisposiciones genéticas igualmente indeseables respecto a las que sí podemos hacer algo. Y es precisamente por este motivo por lo que muchas de las personas que, a la vista de su árbol genealógico, tienen probabilidades de sufrir la enfermedad de Huntington prefieren no hacerse una sencilla prueba que les diría con una certeza prácticamente absoluta si la tienen o no. Pero nótese que tan pronto como se abriera una vía para tratar a aquellos que padecen la mutación de Huntington, cosa que podría suceder en el futuro, esas mismas personas serían las primeras en hacerse la prueba.
Gould y otros han declarado una firme oposición al «determinismo genético», pero dudo que nadie piense que nuestras dotaciones genéticas sean infinitamente revisables. Es prácticamente imposible que yo vaya a dar a luz, dado mi cromosoma Y. Esto es algo que no puedo cambiar ni por voluntad, ni por educación, ni por cultura (al menos no durante mi vida, pero ¿quién sabe lo que hará posible un siglo más de ciencia?). De modo que, al menos por lo que respecta al futuro previsible, algunos de mis genes fijan ciertos aspectos de mi destino sin dejarme ninguna perspectiva real de evadirme de él. Si eso es el determinismo genético, todos somos deterministas genéticos, incluido Gould. En cuanto eliminamos las caricaturas, lo que queda, en el mejor de los casos, son honestas diferencias de opinión respecto al grado de intervención que se requeriría para contrarrestar una u otra tendencia genética y, lo que es más importante, si tal intervención estaría justificada. Se trata de importantes cuestiones morales y políticas, pero muchas veces se hace casi imposible debatir sobre ellas de una forma tranquila y razonable. Un primer paso hacia la recuperación del sentido común es reconocer, como criterio práctico, que, cada vez que nos «acusen» de ser «deterministas genéticos», hay muchas probabilidades de que se trate simplemente de un caso más de ¡Detengan a ese cuervo! y no merece la pena llevar más lejos la discusión, al menos no en esos términos. Por otro lado, ¿qué es lo que hace tan señaladamente perverso el determinismo genético? ¿No sería igual de temible el determinismo del entorno? Consideremos una definición paralela del determinismo del entorno:
Si hemos sido criados y educados en un entorno cultural determinado, entonces los rasgos que tal entorno ha impuesto sobre nosotros son inevitables. Podemos, en el mejor de los casos, canalizarlos de un modo u otro, pero no podemos cambiarlos ni por voluntad, ni por educación ulterior, ni mediante la adopción de una nueva cultura.
A menudo se cita (no sé con qué exactitud) la siguiente frase de los jesuitas: «Dadme un niño hasta los 7 años y yo os devolveré al hombre». Una exageración efectista, sin duda, pero nadie duda de que la educación temprana y otros eventos importantes de la infancia pueden tener un efecto importante sobre la vida posterior. Hay estudios, por ejemplo, que sugieren que ser rechazado por la madre en el primer año de vida o ser víctima de otros hechos traumáticos de este tipo aumentan la probabilidad de cometer un crimen violento (por ejemplo, Raine y otros, 1994). De nuevo, no debemos caer en el error de equiparar el determinismo con la inevitabilidad. Lo que debemos examinar empíricamente —y es algo que varía mucho tanto en el plano del entorno como en el genético— es si los efectos indeseados, por más grandes o profundos que puedan ser, son evitables con sólo tomar ciertas medidas. Consideremos el mal conocido como no saber una palabra de chino. Yo personalmente lo padezco, totalmente por causa de influencias del entorno durante mi infancia temprana (mis genes no tenían nada que ver con ello, al menos directamente). Si tuviera que mudarme a China, sin embargo, pronto estaría «curado», sólo con que pusiera algo de esfuerzo de mi parte, aunque sin duda conservaría signos profundos e insuperables de mi deficiencia, fácilmente detectables para cualquier hablante nativo de chino, durante el resto de mi vida. Pero ciertamente conseguiría arreglármelas lo bastante bien en chino como para ser responsable de las acciones que pudiera emprender bajo la influencia de los hablantes de chino que me encontrara.
¿Acaso no es cierto que todo lo que no viene determinado por nuestros genes debe venir determinado por nuestro entorno? ¿Qué más puede haber? Está la Naturaleza y está la Crianza. ¿Hay alguna otra X, algún factor ulterior que contribuya a lo que somos? Está el Azar. La Suerte. Ya hemos visto en los capítulos 3 y 4 que este ingrediente extra es importante, pero no tiene por qué venir de las entrañas cuánticas de nuestros átomos o de ninguna estrella lejana. Nos rodea por todas partes en los eventos azarosos y carentes de causa de nuestro ruidoso mundo, que llenan automáticamente todas las lagunas de especificación que dejan sin fijar nuestros genes o las causas más prominentes que operan en nuestro entorno. Esto es particularmente evidente en la formación de los billones de conexiones en nuestro cerebro. Hace años que se sabe que el genoma humano, a pesar de su extensión, es con mucho demasiado pequeño para especificar (en su receta genética) todas las conexiones que se forman entre las neuronas. Lo que ocurre es que los genes especifican procesos que disparan grandes aumentos en la población de neuronas —muchas más de las que nuestros cerebros usarán nunca—, las cuales despliegan terminaciones de manera aleatoria (pseudoaleatoria, por supuesto) hasta que en muchos casos conectan casualmente con otras neuronas de formas que resultan útiles en un sentido detectable[3](detectable para los inconscientes procesos de poda del cerebro). Estas conexiones ganadoras tienden a sobrevivir, mientras que las conexiones perdedoras mueren, para ser luego desmanteladas de modo que sus partes puedan reciclarse unos días más tarde en la próxima hornada de crecimientos neuronales. Este entorno selectivo en el interior del cerebro (especialmente en el cerebro del feto, mucho antes de que llegue al entorno exterior) no especifica las conexiones finales más de lo que puedan hacerlo los genes; hay factores, tanto en los genes como en el entorno, que influyen y podan su crecimiento, pero hay mucho que queda en manos del azar.
Cuando recientemente se publicó el genoma humano y se anunció que «sólo» tenemos alrededor de 30 000 genes (según los criterios actuales sobre cómo identificar y contabilizar los genes), no los 100 000 que habían estimado algunos expertos, corrió un divertido suspiro de alivio entre la prensa. ¡Uf! No somos meros productos de nuestros genes: ¡«nosotros» aportamos todas las especificaciones que de otro modo habrían «fijado» aquellos 70 000 genes! Pero cabe preguntarse: ¿cómo vamos a cumplir nosotros con esta tarea? ¿No nos exponemos a una amenaza igual por parte de nuestro temible entorno, de la vieja Crianza con sus insidiosas técnicas de adoctrinamiento? ¿Cuando la Naturaleza y la Crianza hayan hecho su trabajo, quedará algo para que yo pueda ser jo? (Si uno se hace realmente pequeño, puede externalizarlo prácticamente todo).
¿Acaso importa cuál sea el compromiso concreto si, sea como sea, son nuestros genes y nuestro entorno (incluido el azar) los que se reparten el botín y «fijan» nuestros caracteres? Tal vez parezca que el entorno es una fuente más benigna de determinación, pues, después de todo, «podemos cambiar el entorno». Eso es cierto, pero no podemos cambiar el entorno pasado de una persona más de lo que podemos cambiar a sus padres, y los ajustes futuros en el entorno pueden dirigirse con igual firmeza a compensar limitaciones genéticas previas como limitaciones ambientales previas. Y en la actualidad estamos a un paso de poder ajustar el futuro genético casi con la misma facilidad que el futuro entorno. Supongamos que sabemos que uno de nuestros hijos tendrá un problema que puede mitigarse con un reajuste en sus genes o bien en su entorno. Puede haber muchas razones válidas para preferir un tratamiento que otro, pero ciertamente no es evidente que una de esas opciones deba ser descartada sobre bases morales o metafísicas. Suponga, por proponer un caso imaginario que probablemente se verá superado pronto por la realidad, que usted es un inuit convencido para quien la vida por encima del Círculo Ártico es la única que vale la pena vivir, y supongamos que le dicen que sus hijos estarán genéticamente incapacitados para vivir en un entorno de este tipo. Puede mudarse a los trópicos, donde sus hijos estarán bien —al precio de renunciar a su herencia ambiental—, o puede reajustar su genoma para que puedan seguir viviendo en el mundo Ártico, al precio (si es que puede considerarse así) de la pérdida de algún aspecto de su herencia genética «natural».
La cuestión no es el determinismo, sea genético o del entorno, o de ambos a la vez; la cuestión es qué podemos cambiar, sea o no determinista el mundo. Jared Diamond ofrece una fascinante perspectiva sobre la equívoca cuestión del determinismo genético en su magnífico libro Armas, gérmenes y acero (1997). La cuestión que plantea Diamond, y que en buena medida responde, es por qué los «occidentales» (los europeos o los euroasiáticos) han logrado conquistar, colonizar y en sentido amplio dominar a la gente del «Tercer Mundo», y no ha sucedido al revés. ¿Por qué las poblaciones humanas de América o África, por ejemplo, no crearon imperios mundiales capaces de invadir, masacrar y someter a los europeos? ¿Es la respuesta… la genética? ¿Acaso ha demostrado la ciencia que la fuente última del dominio de Occidente reside en nuestros genes? Cuando ven planteada por primera vez esta pregunta, muchas personas —incluso científicos muy competentes— dan por supuesto que Diamond, por el mero hecho de plantearla, debe suscribir alguna hipótesis racista acerca de la superioridad genética de los europeos. Tanto les inquieta su sospecha que les cuesta mucho comprender que lo que dice Diamond (y debe esforzarse mucho para dejarlo claro) viene a ser lo contrario: la explicación secreta no reside en nuestros genes, los genes humanos, pero sí en buena medida en otros genes: los genes de las plantas y los animales que fueron los antepasados silvestres de todas las especies domesticadas que se emplean en la agricultura humana.
Los guardias de prisión tienen un dicho: si algo puede ocurrir, ocurrirá. Quieren decir con ello que si existe algún fallo en la seguridad, alguna prohibición o vigilancia inefectivas o algún punto débil en las barreras, pronto serán encontrados y explotados hasta donde sea posible por los prisioneros. ¿Por qué? La perspectiva intencional lo deja bien claro: los prisioneros son sistemas intencionales frustrados que se caracterizan por la inteligencia y la astucia; eso significa mucho deseo informado con una gran cantidad de tiempo libre para explorar su entorno. Su procedimiento de búsqueda será tan bueno como exhaustivo, y sabrán distinguir las mejores estrategias de las que no lo son tanto. No hay duda de que encontrarán cualquier cosa que se pueda encontrar. Diamond parte del mismo criterio, y supone que la gente de cualquier lugar del mundo ha sido siempre más o menos igual de lista, eficiente y oportunista, igual de disciplinada, previsora, como la de cualquier otra parte del mundo, y, por lo tanto, que la gente ha encontrado siempre lo que había por encontrar. En una primera aproximación, puede decirse que todas las especies salvajes domesticabas han sido domesticadas. La razón por la que los euroasiáticos tomaron la delantera en el terreno tecnológico fue porque tomaron la delantera en el terreno agrícola, y lo consiguieron porque entre las plantas y los animales salvajes que tenían a su alrededor hace diez mil años había candidatos ideales para la domesticación. Había hierbas que eran genéticamente fáciles de convertir en superplantas: sólo hacían falta unas pocas mutaciones para que pasaran a ser nutritivos y generosos cereales, algo que podía conseguirse más o menos por accidente; también había animales que por su naturaleza social estaban genéticamente cerca de convertirse en animales de pastoreo y que se reproducían fácilmente en cautividad. (El maíz costó más de domesticar en el hemisferio occidental en parte porque debía recorrer una mayor distancia genética respecto a su precursor silvestre.) La mayor parte de los factores de selección que permitieron cubrir este camino, antes de la llegada de la agronomía moderna, correspondieron sin duda a lo que Darwin llamaba la «selección inconsciente»: las tendencias en gran medida involuntarias y ciertamente no informadas que estaban implícitas en los patrones de comportamiento de unas personas que tenían una perspectiva tremendamente limitada sobre lo que hacían y por qué lo hacían. Las causas principales, los factores que «fijaron» las oportunidades de la gente en cada lugar, fueron ante todo los accidentes biogeográficos y, por lo tanto, el entorno. Gracias a haber vivido durante milenios rodeados de muchas variedades de animales domesticados, los eurasiáticos desarrollaron además inmunidad hacia diversos agentes patógenos susceptibles de transmisión entre animales y humanos —aquí sí que hay que destacar un papel importante que corresponde a los genes humanos, confirmado más allá de cualquier sombra de duda— y cuando, gracias a su tecnología estuvieron en condiciones de recorrer largas distancias y encontrarse con otros pueblos, sus gérmenes hicieron muchas veces tanto daño como sus armas y su acero.
¿Qué podemos decir de la tesis de Diamond? ¿Es un peligroso determinista genético, o un peligroso determinista ambiental? No es ninguna de las dos cosas, naturalmente, pues las dos modalidades de hombre del saco son tan míticas como los hombres-lobo. Al aumentar la información que tenemos a nuestro alcance acerca de las diversas causas que llevaron a las condiciones que limitan nuestras oportunidades actuales, ha aumentado nuestra capacidad para evitar y prevenir lo que queramos prevenir. El conocimiento del rol que desempeñan nuestros genes y los genes de las especies que nos rodean no es un enemigo de la libertad humana, sino uno de sus mejores aliados.
Para poder percibir las «decisiones» que toman los linajes (sean de células parásitas o sean de secuoyas) hay que mirar de la manera adecuada. Es necesario adoptar la perspectiva intencional hacia esas curiosas agregaciones de materia, rebobinar el tiempo hacia adelante y esperar a que los patrones de alto nivel emerjan, como en efecto lo hacen, en medio de la montaña de datos con agradable predictibilidad. Las decisiones más reconocibles, realizadas en tiempo real por individuos compactos y fácilmente identificables, tuvieron que esperar al surgimiento de la locomoción. Sí, los árboles pueden «decidir» que la primavera ha llegado y que es hora de hacer brotar sus flores, y las almejas pueden «decidir» cerrarse cuando perciben un golpe alarmante sobre sus conchas, pero esas elecciones son tan rudimentarias, tan parecidas al funcionamiento de un simple interruptor, que sólo las consideramos decisiones por cortesía. Pero incluso los interruptores, que se limitan a encenderse o apagarse como consecuencia de ciertos cambios en el entorno, dan lugar a un cierto grado de libertad, tal como dicen los ingenieros, y requieren por lo tanto un cierto grado de control, sea del tipo que sea. Un sistema posee un cierto grado de libertad cuando hay un conjunto de posibilidades diferentes y la actualización de una u otra de estas posibilidades depende de la función o del interruptor que controla esta libertad. Los interruptores (que pueden tener sólo dos o bien múltiples posiciones) pueden estar conectados unos a otros en serie, en paralelo o en sistemas que combinen ambos tipos de conexión. A medida que estos sistemas proliferan, y forman redes de interruptores cada vez más grandes, los grados de libertad se multiplican de forma asombrosa y los problemas de control se vuelven complejos y no lineales. Cualquier linaje equipado con una estructura de este tipo se enfrenta a un problema: ¿qué información debería modular el paso por esta red de caminos que se bifurcan en un espacio multidimensional de posibilidades? Para eso está el cerebro.
Un cerebro, con sus bancos de inputs sensoriales y outputs motores, es un dispositivo centralizado para rastrear el entorno pretérito en busca de información que pueda luego refinar hasta dar con las pepitas de oro de unas expectativas válidas sobre el futuro. Luego cada uno puede usar estas expectativas que tanto trabajo le ha costado conseguir para modular sus elecciones, mejor de lo que sus congéneres modulan las suyas. La velocidad es esencial, puesto que el entorno cambia constantemente y está plagado de competidores, pero también lo es la precisión (puesto que una de las opciones de los competidores es el camuflaje) y, por lo tanto, la eficiencia (puesto que todo tiene un precio y al final tiene que dar resultados). Estas condiciones evolutivas dan lugar a un conjunto de compromisos que premian una atención sensorial rápida, de alta precisión y muy focalizada. La carrera armamentística garantiza que cada especie ignorará tantos aspectos como pueda de su entorno, una estrategia peligrosa que puede dar lugar a sorpresas desagradables en el futuro, cuando una variable del entorno que hasta el momento había resultado trivial cobre de repente una relevancia crucial.
Esta perspectiva de orden superior sobre un entorno rico en novedades no previsibles y sin embargo relevantes plantea otra posible apuesta: ¿saldrá a cuenta que algún linaje invierta en aprendizaje? Dicha apuesta tiene un coste sustancial de carácter general: es preciso instalar cierta maquinaria para hacer posibles unas redes de interruptores que puedan rediseñarse en tiempo real, durante la vida del propio individuo, para que este pueda ajustar sus funciones de control en respuesta a las nuevas pautas que detecte en el mundo. Recordemos la distinción de Drescher (1991) entre las máquinas de situación-acción y las máquinas de elección mencionada en el capítulo 2. Las máquinas de situación-acción consisten en un sistema de interruptores relativamente sencillos, cada uno de los cuales encarna lo que viene a ser una especie de regla de respuesta al entorno: si encuentras la condición C, haz Estos interruptores son eficientes para organismos relativamente simples cuyo comportamiento está especificado desde el nacimiento. Las máquinas de elección tienen un conjunto de mecanismos distintos, que encarnan predicciones del tipo: si encuentras la condición C, hacer A tendría el resultado Z (con la probabilidad p). Generan varias o muchas predicciones de este tipo, y luego las evalúan (usando cualesquiera valores que tengan previamente o hayan desarrollado ellos mismos), un funcionamiento que resulta eficiente para organismos diseñados para aprender en el curso de su vida. Un organismo puede tener instaladas ambas clases de maquinaria y confiar en la primera para realizar elecciones rápidas y poco refinadas para salvar la vida, y en la última para pensar seriamente acerca del futuro: una rudimentaria facultad de razonamiento práctico.
Esta sofisticada maquinaria para el aprendizaje sólo saldrá a cuenta si hay suficientes ocasiones para aprender (y si el aprendizaje tiende a ir en la dirección de adquirir hábitos buenos, no hábitos malos, por supuesto). ¿De cuántas ocasiones estamos hablando? Eso depende de las circunstancias, pero no hay duda de que a menudo no las hay. «Úsalo o piérdelo» es un lema que tiene muchas aplicaciones en el mundo animal. Por ejemplo, los cerebros de los animales domesticados son significativamente más pequeños que los cerebros de sus parientes más próximos en estado salvaje, y eso no es sólo un producto secundario de la selección en busca de una mayor masa muscular en animales criados para buscarse su propio alimento. Los animales domesticados pueden permitirse ser estúpidos sin dejar de tener una descendencia muy numerosa, pues lo que han hecho ha sido delegar muchas de sus tareas cognitivas en otra especie, nosotros, de la que se han convertido en parásitos. Del mismo modo que las tenías han «decidido» confiar en nosotros para realizar todas sus tareas de locomoción y búsqueda de alimento, gracias a lo cual pueden simplificar drásticamente unos sistemas nerviosos que ya no necesitan, los animales domesticados se verían en dificultades si no pudieran contar con sus anfitriones humanos. No son entoparásitos que vivan dentro de nosotros, pero siguen siendo parásitos.
Estamos ya muy cerca de la libertad del pájaro, que puede volar e ir allí donde quiera. ¿Por qué quiere el pájaro ir allí donde quiera? Tiene sus motivos. Sus razones están encarnadas en la configuración de los interruptores de su cerebro, y vienen avaladas, a largo plazo, por su continuada supervivencia. Principalmente, las cosas sobre las que el pájaro se toma la molestia de reunir información son las más relevantes para su bienestar inmediato. Cuanta más presión de algún taimado competidor hayan sufrido sus ancestros en los últimos tiempos, más probable es que lleve instalado un costoso equipo para contrarrestar sus amenazas. Cuando los marineros llegaron por primera vez con sus barcos a las remotas islas del Pacífico, habitadas por pájaros cuyos ancestros llevaban miles de años sin ver a ningún depredador, encontraron ejemplares tan poco curiosos, tan poco temerosos de las grandes cosas en movimiento que se les acercaban, que los marineros podían tender la mano y cogerlos. Esos pájaros eran perfectamente capaces de volar, pero no hacía falta tomar la menor precaución para capturarlos. Podían volar y desplazarse hasta donde quisieran, pero les faltaba picardía; había razones en el aire para hacer ciertas cosas, pero ellos no eran lo bastante listos como para captarlas. Tenían gran cantidad de oportunidades en bruto de salvarse, pero les faltaba la información necesaria para hacerlas efectivas. Por supuesto, la mayor parte de esas especies de pájaros están hoy extinguidas.
La carrera armamentística entre el depredador y la presa, así como la competición entre congéneres por el apareamiento y por los medios que llevan al apareamiento —comida, refugio, territorio, posición dentro del grupo, etc.—, han dado lugar en nuestra biosfera a cientos de miles de años de I+D en un amplio espectro de procesos paralelos y simultáneos protagonizados por millones de especies. En este mismo instante, billones de organismos de este planeta están jugando al escondite. Pero para ellos no es sólo un juego; es una cuestión de vida o muerte. Hacer/o bien, no cometer errores, es importante para ellos —en realidad no hay nada más importante—, pero en general no son conscientes de ello. Son los beneficiarios de un equipo exquisitamente diseñado para hacer bien lo que deben hacer, pero cuando este equipo no funciona como es debido y lo hace mal, no disponen de ningún recurso, por lo general, para darse cuenta de ello, y ya no digamos para lamentarlo. Siguen adelante, sin darse cuenta del problema. La diferencia entre lo que parecen las cosas y lo que son realmente es una diferencia tan crucial para ellos como pueda serlo para nosotros, pero les resulta en buena medida desconocida. El reconocimiento de la diferencia entre la apariencia y la realidad es un descubrimiento humano. Sólo unas pocas especies —algunos primates, algunos cetáceos, tal vez incluso algunos pájaros— muestran signos de apreciar el fenómeno de la «falsa creencia» (hacerlo mal). Muestran sensibilidad a los errores de otros en cuanto errores, y tal vez incluso cierta sensibilidad hacia los suyos propios, pero carecen de la capacidad de reflexión necesaria para concentrarse en esta posibilidad y, por lo tanto, no pueden usar esta sensibilidad para el diseño deliberado de reparaciones o mejoras en su propio equipo de búsqueda o de ocultación. Esa brecha entre la apariencia vía realidad es algo que sólo los seres humanos hemos logrado salvar.
Somos la especie que descubrió la duda. ¿Hay bastante comida almacenada para el invierno? ¿He calculado mal? ¿Me está engañando mi pareja? ¿Debería haberme desplazado hacia el sur? ¿Es seguro entrar en esta cueva? Otras criaturas se muestran a veces visiblemente agitadas por sus propias incertidumbres respecto a cuestiones parecidas, pero como no pueden hacerse a sí mismas esas preguntas, no pueden articular sus problemas por sí mismas o dar pasos para acercarse más a la verdad. Están atrapadas en un mundo de apariencias, sacan tanto partido como pueden a lo que parecen ser las cosas y raras veces se preocupan, si es que llegan a hacerlo nunca, por si son verdaderamente tal como parecen. Sólo a nosotros pueden asaltarnos las dudas, y sólo a nosotros nos ha aguijoneado esta inquietud epistémica para buscar formas de remediarla: mejores métodos para la búsqueda de la verdad. En nuestro intento de mejorar el control que tenemos sobre nuestras fuentes de alimento, nuestros territorios, nuestras familias, nuestros enemigos, descubrimos los beneficios de hablar con los demás, hacer preguntas, transmitir la tradición. Inventamos la cultura.
Es la cultura lo que nos da el punto de apoyo que nos permite elevarnos a un territorio nuevo. La cultura nos da una atalaya desde la cual podemos ver cómo cambiarlas trayectorias hacia el futuro que han diseñado las investigaciones ciegas de nuestros genes. Tal como ha dicho Richard Dawkins: «Lo importante es que no existe ninguna razón general para suponer que las influencias genéticas sean más irreversibles que las del entorno» (Dawkins, 1982, pág. 13). Pero para revertir esta clase de influencias, debemos ser capaces de reconocerlas y comprenderlas. Sólo nosotros, los seres humanos, disponemos del conocimiento a largo plazo necesario para identificar y evitar las trampas que se esconden en los caminos proyectados por nuestros genes carentes de previsión. El conocimiento compartido es la clave para aumentar nuestra libertad respecto al «determinismo genético».
No hemos llegado aún a la sala de conciertos, pero estamos cada vez más cerca.
Capítulo 5
La sabiduría implícita en el diseño de las formas de vida multicelulares sólo se entiende adecuadamente cuando se adopta la perspectiva intencional respecto al conjunto del proceso evolutivo. Desde esta perspectiva podemos discernir las rabones virtuales que hay detrás de las «elecciones» cooperativas en juegos de no suma cero que han guiado los procesos evolutivos de I+D hacia la creación de agentes racionales cada vez más sofisticados, y hacia la multiplicación de la capacidad de las formas de vida para reconocer y reaccionar ante las oportunidades. Cuando nos olvidamos del espantajo del «determinismo genético», vemos como la evolución guiada por la selección natural hace posibles unos niveles cada vez mayores de libertad, aunque no se trata aún de la libertad de la agencia humana.
Capítulo 6
La cultura humana no es ni un milagro ni una más de las herramientas que nos proporcionan nuestros genes para mejorar su propia competencia. Para comprender cómo es posible que una persona sea a la vez creadora de cultura y una creación de la misma debemos examinar el proceso evolutivo que ha llevado a lo largo de múltiples etapas al surgimiento de la cultura y la sociabilidad humana.
Pueden encontrarse desarrollos más extensos de las ideas presentadas en este capítulo en La peligrosa idea de Darwin (Dennett, 1995), de donde proceden algunos párrafos del capítulo Games, Sex and Evolution, de John Maynard Smith (1988; sobre todo los capítulos 21 y 22) es una excelente introducción a la aplicación de la teoría de juegos a la evolución, igual que la edición revisada de El gen egoísta (1976), de Richard Dawkins. Evolution of the Social Contract (1996), de Brian Skyrms, pasa revista a investigaciones más recientes. Para un atractivo repaso general a la perspectiva esbozada en este capítulo, véase Nonzero: The Logic of Human Destiny (2000), de Robert Wright.
Nuestra comprensión de los procesos evolutivos aquí descritos, en especial de los conflictos entre genes que pueden describirse desde una perspectiva intencional, aumenta con gran rapidez. Muchas de las tesis específicas que se sostienen hoy (como el número de genes del genoma humano) puede que queden canceladas mañana, pero el esqueleto de la teoría y la evidencia que está en la base de la biología evolutiva es notablemente firme y resistente. Un libro excelente, aunque difícil, que pasa revista a los pasos en la transición desde las formas de vida más sencillas hasta las sociedades humanas es The Major Transitions in Evolution (1995), de Maynard Smith y Eórs Szathmáry; una versión más sencilla es su libro de 1999, Ocho hitos de la evolución: del origen de la vida a la aparición del lenguaje. Para una revisión autorizada del estado de la cuestión hacia finales de 2000, véase Evolution: From Molecules to Ecosystems, editado por Andrés Moya y Enrique Font (de próxima aparición), donde se recogen una serie de estudios sobre temas tales como la evolución de la multicelularidad, los conflictos que pueden surgir a pesar del destino en buena medida compartido de los genes mitocondriales y nucleares, de los compromisos coste-beneficio de la simbiosis, y muchos otros temas fascinantes.
La distinción de Drescher entre las máquinas de situación-acción y las máquinas de elección clarifica de forma útil la distinción (y en parte coincide con ella) que propongo entre criaturas skinnerianas y popperianas (Dennett, 1975, 1995, 1996a).