A
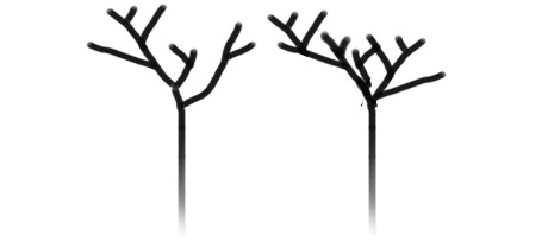
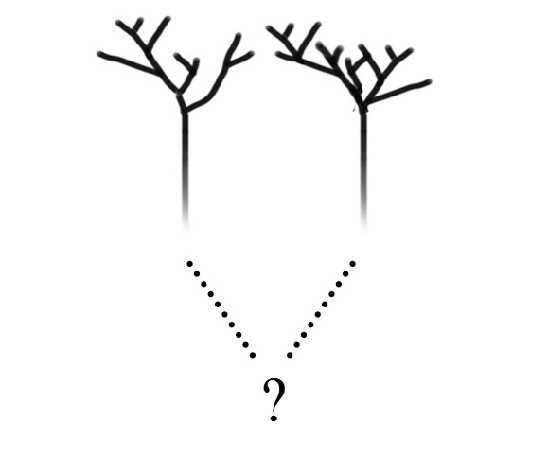
Si algún día llegamos a descubrir vida extraña, la prioridad será determinar si pertenece a un árbol de la vida verdaderamente distinto o si, por el contrario, no es más que una rama del árbol de la vida conocido que todavía no habíamos descubierto. Esta distinción se ilustra en la figura 4. Supongamos que nos presentan dos formas de vida radicalmente distintas, que estamos tentados a atribuir a árboles separados, cada uno de ellos con un origen independiente (con lo que me refiero a transiciones independientes de la materia inerte a la vida), tal como se muestra en la figura 4a. Sin embargo, al investigarlas más a fondo podríamos descubrir que «bajo tierra» las dos ramas se unen en un sistema radical común (figura 4b), es decir, que en último análisis las formas de vida distintas pertenecen a un mismo árbol, pero divergieron antes que el último antepasado común de la vida estándar.
El árbol de la vida que conocemos consiste en tres «ramales» diferenciados que se separaron hace miles de millones de años (véase la figura 2). Uno de los ramales contiene las bacterias. Otro contiene toda la vida pluricelular, desde los humanos a los erizos. También incluye organismos unicelulares complejos como las amebas. Éste es el dominio de los eucariotas o «Eucarya». El tercer ramal está formado únicamente por microbios, pero tan diferentes de las bacterias como éstas lo son de nosotros, y reciben el nombre colectivo de arqueas o «Archaea». La cuestión que planteo es: ¿cómo sabemos que no existe un cuarto ramal, todavía por descubrir, que se separó aún antes de la trifurcación en bacterias, eucariotas y arqueas? Si alguna vez descubriéramos una forma de vida exótica, tendríamos que eliminar la explicación del «cuarto ramal» antes de concluir que constituye una prueba de un segundo árbol.
¿Cómo puede distinguirse una rama baja y enterrada de un árbol distinto? La respuesta dependerá en parte de lo extraña que sea la vida extraña. Por decirlo con una frase manida, el diablo está en los detalles. Pensemos en el caso de la «vida especular» (organismos con la quiralidad invertida). ¿Es concebible que las primeras formas de vida fuesen aquirales, es decir, construidas sólo a partir de moléculas con simetría especular, tras lo cual la vida se hubiera dividido en dos dominios, uno poblado por organismos con azúcares levógiros y aminoácidos dextrógiros, y el otro por organismos con las formas especulares? Parece extremadamente improbable. Las moléculas pequeñas y simples suelen mostrar simetría especular, pero las moléculas de complejidad siquiera moderada tienen versiones levógira y dextrógira. Es muy difícil concebir que un sistema con la complejidad de un organismo vivo pueda construirse únicamente con moléculas aquirales. Así que el descubrimiento de vida especular sería una fuerte indicación de acontecimientos múltiples de génesis.
A
B
FIGURA 4. ¿Árbol o bosque? Si en la Tierra coexistieran dos formas de vida, sería importante determinar si corresponden a árboles de la vida separados, con origen independiente, como en A, o si simplemente a lo largo de su evolución han divergido mucho a partir de un único origen común, como en B.
En cambio, si se descubriese una forma de vida extraña que se pareciera a la vida estándar en todo menos en el código genético sería fácil defender la tesis de que ambas formas tuvieron una génesis común y un código precursor común, tras lo cual la vida se habría dividido en dos formas que evolucionaron hacia códigos distintos. Al menos una versión de este escenario parece plausible. El código de tripletes que utiliza la vida estándar es complicado, y algunos biólogos han conjeturado que evolucionó a partir de un precursor más sencillo, tal vez un código de dobletes basado en sólo dos nucleótidos (G y C) y diez aminoácidos. Cabría esperar que esta versión reducida de la vida estándar fuese menos compleja, pero tal vez pudiera prosperar perfectamente hace tres o cuatro mil millones de años. El código de tripletes podría haber evolucionado más tarde, ofreciendo una mayor versatilidad que habría permitido a la vida expandirse en un abanico aún más amplio de ambientes. La transición de un código de dobletes a uno de tripletes podría haberse producido más de una vez, o el código de tripletes original se podría haber dividido con posterioridad en variantes.
Pero se presenta una posibilidad todavía más interesante. ¿Es posible que algunos de esos «tipos de antaño» anden todavía por ahí, viviendo al viejo estilo de un código de dobletes G-C? Una vez más, estos «fósiles vivientes» se pasarían por alto en los análisis bioquímicos estándar, pero serían fáciles de identificar si los investigadores decidieran buscarlos.[3.20] Del mismo modo, si se descubriese vida que usase arsénico, tendríamos que averiguar si la vida estándar comenzó de ese modo y luego evolucionó reemplazando el arsénico con el fósforo. Pero por fascinante que pueda ser el descubrimiento de tales organismos precursores, no llegaría al verdadero núcleo de la cuestión, que es la posibilidad de orígenes múltiples. Para estar seguros de que una vida extraña desciende realmente de una segunda génesis, tendría que ser lo bastante distinta de la vida estándar para que no pudiera haber existido un antepasado común plausible. Ese criterio sería difícil de establecer si las dos biosferas se solaparan y utilizaran mucha química en común. Aún más difícil sería si las dos formas llegaran a estar parcialmente integradas bioquímicamente, por ejemplo, mediante el intercambio de genes u otras estructuras, haciendo de este modo borrosa la división entre sus linajes separados y confusa toda su historia evolutiva. No podemos descartar que una forma de vida «ocupase» a otra, al estilo de Quatermass, infiltrando componentes básicos de sí misma en un hospedador receptivo, sobre todo si las dos formas separadas de vida se encuentran en vías evolutivas convergentes. Todo esto sería una complicación indeseable. Sería triste y enojoso que la vida hubiera comenzado en la Tierra muchas veces, pero hubiera convergido y se hubiera mezclado, dejándonos sin esperanzas de desenmarañar sus múltiples raíces.[3.21] Personalmente, no creo que la convergencia evolutiva pueda llegar a ser tan fuerte. Podría producir grandes rasgos que se asemejen, pero parece poco probable que pueda implicar esquemas bioquímicos específicos.
Suele argumentarse que si dos formas de vida distintas se encontraran codo con codo, una de ellas acabaría por obtener una ventaja y eliminar a la otra. Nunca he llegado a convencerme de que las cosas hayan de desarrollarse así. La existencia pacífica es otra posibilidad; puede surgir de dos modos. En primer lugar, si las dos formas son lo bastante distintas como para convivir con total indiferencia mutua, no competirían de ningún modo. Por ejemplo, la vida especular no competiría directamente con la vida conocida porque, en gran medida, estas dos formas de vida utilizarían moléculas distintas como alimento. Una de las formas podría llegar a dominar en términos numéricos, pero ¿y qué? Los microbiólogos están familiarizados con el hecho de que algunas especies son muy raras, pese a lo cual constituyen un componente estable de la población microbiana total. El segundo tipo de coexistencia pacífica se produce cuando las poblaciones de tipos microbianos muy dispares alcanzan alguna forma de acomodo. Encontramos un ejemplo en la convivencia codo con codo de bacterias y arqueas, dos grandes dominios microbianos que comprenden millones de especies que a menudo comparten nichos parecidos. Puede pensarse que esta tolerancia se debe a que los dos dominios han alcanzado algún tipo de integración bioquímica, matrimonio en lugar de rivalidad. El intercambio de genes se produce continuamente en la vida, sobre todo en los microbios. Pero, en realidad, las arqueas y las bacterias han protegido celosamente ciertos genes fundamentales. Por lo que sabemos, las arqueas nunca han compartido con las bacterias (o los eucariotas) su capacidad para realizar un metabolismo que produce metano; sin embargo, la metanogénesis es común en las arqueas, y se produce en lugares tan diversos como las chimeneas del océano profundo y el intestino humano. En el sentido contrario, no parece que la fotosíntesis haya pasado nunca de las bacterias (o los eucariotas) a las arqueas.[3.22] Así que es evidente que formas muy distintas de microbios pueden competir en el mismo espacio por muchos recursos comunes, sin que una forma acabe por eliminar a la otra.
Incluso si los descendientes de otros orígenes se extinguieron hace mucho tiempo, tal vez hayan dejado algún resto de su antigua existencia en forma de fósiles o biomarcadores moleculares distintivos. Por ejemplo, los esteranos (moléculas con cuatro anillos) son producidos por células complejas y, que se sepa, no pueden formarse por ningún medio abiótico. Los esteranos se han encontrado en trazas en microfósiles que datan de hace 2.700 millones de años. Si se descubrieran fósiles que contuvieran esteranos «especulares», o sea, con la quiralidad opuesta, podría considerarse un indicio de vida especular en el pasado. Muchas otras moléculas orgánicas complejas producidas por vías metabólicas radicalmente distintas podrían sobrevivir en rocas durante mucho tiempo. Una forma indirecta en que la vida extraña extinta podría haber dejado una traza es a través de su acción sobre minerales. Se cree que muchos depósitos minerales, entre ellos los de hierro, cobre y oro, son biogénicos, es decir, su deposición y concentración fueron causadas, al menos en parte, por la actividad de microbios que utilizaban esos metales en su metabolismo. El hallazgo de un depósito mineral que no haya podido crear la vida conocida, pero presentara los signos de un origen biogénico, sería una prueba circunstancial del funcionamiento de una bioquímica alternativa.