FIGURA 8.1. Vista desde el multiverso de dos segmentos de ADN que resultan ser idénticos en nuestro universo, uno aleatorio y el otro del interior de un gen.
Desde la antigüedad hasta, aproximadamente, el siglo XIX se daba por sentado que era necesaria la existencia de una fuerza o factor animante especial para que la materia de los organismos vivos se comportara de modo tan notoriamente diferente que el resto de la materia. Se postulaba, pues, la existencia de dos clases de materia, animada e inanimada, dotadas de propiedades físicas distintas por completo. Consideremos un organismo vivo, por ejemplo un oso. Una fotografía de un oso se parece a un oso en determinados aspectos. También se parecen a un oso otros objetos inanimados, tales como el cadáver de un oso e incluso, de un modo más limitado, la constelación de la Osa Mayor. Pero sólo la materia animada puede perseguirnos a través del bosque mientras intentamos escapar zigzagueando entre los árboles y destrozarnos si nos atrapa. Las cosas inanimadas nunca harían algo tan premeditado… o, al menos, así lo creían los antiguos. Por supuesto, no habían visto nunca un cohete dirigido.
Para Aristóteles y otros filósofos de la antigüedad, la facultad más espectacular de la materia animada era su capacidad de moverse voluntariamente. Consideraban que cuando la materia inanimada, como una piedra, queda en reposo, no vuelve a moverse a menos de ser golpeada por algo. La materia animada, en cambio, puede estar en reposo, como un oso en hibernación, y reanudar luego el movimiento sin necesidad de ser golpeada. Gracias a la ciencia moderna, podemos fácilmente detectar fallos en estas generalizaciones. Por ejemplo, la idea de «movimiento voluntario» parece ahora errónea: sabemos que el oso se despierta a causa de los procesos electroquímicos de su cuerpo. Éstos pueden ser iniciados por «golpes» externos, tales como cambios de temperatura, o por su reloj biológico interno, que utiliza lentas reacciones químicas para seguir el paso del tiempo.
Las reacciones químicas son movimientos de átomos, de modo que el oso nunca se encuentra en completo reposo. Por otro lado, un núcleo de uranio, que, sin duda, no está vivo, puede permanecer estable durante miles de millones de años hasta que, sin ningún estímulo exterior, un buen día se desintegra de manera súbita y violenta. Así pues, el contenido nominal de la idea de Aristóteles no es válido en la actualidad. No obstante, el Estagirita tenía muy claro algo importante, en lo que, en cambio, se equivocan muchos de los modernos pensadores. Al tratar de asociar la vida con un concepto físico básico (aunque fuera el de movimiento, en lo que estaba equivocado), reconocía que ésta es un fenómeno fundamental de la naturaleza.
Un fenómeno es «fundamental» cuando de su comprensión depende que se comprenda de manera más profunda el mundo. Por supuesto, no existe unanimidad acerca de qué aspectos del mundo vale la pena comprender y, por consiguiente, acerca de lo que es profundo o fundamental. Algunos dirán que el amor es el fenómeno más fundamental del mundo. Otros piensan que, si se aprenden ciertos textos sagrados de memoria, se entiende todo lo que vale la pena entender. La comprensión de la que hablo se expresa en leyes de física y en principios de lógica y filosofía. Una comprensión «más profunda» es aquella que resulta más general, implica menos conexiones entre verdades superficialmente diversas, explica más y deja menos asunciones sin explicar. Los fenómenos más fundamentales intervienen en la explicación de muchísimos fenómenos, pero son explicados únicamente por leyes y principios básicos.
Al contrario de lo que ocurre con la gravitación, que es de los más fundamentales, no todos los fenómenos de esta clase tienen grandes efectos físicos. Los efectos directos de la interferencia cuántica, como las conformaciones fantasmas descritas en el capítulo 2, son tan reducidos que resulta incluso difícil su detección inequívoca. Sin embargo, hemos visto que la interferencia cuántica es un fenómeno fundamental. Sólo si lo comprendemos podremos entender el hecho básico acerca de la realidad física, es decir, la existencia de universos paralelos.
Resultaba evidente para Aristóteles que la vida es fundamental teoréticamente y tiene importantes efectos físicos. Como veremos, tenía razón, pero ello resultaba obvio para él por razones erróneas; por ejemplo, las supuestamente distintas propiedades mecánicas de la materia animada y la dominación de la superficie de la Tierra por los procesos vitales. Aristóteles pensaba que el universo consistía, básicamente, en lo que ahora denominamos la biosfera (la región que contiene la vida) de nuestro planeta, con algunas regiones adicionales (las esferas celestiales y el interior de la Tierra) clavadas, por así decirlo, encima y debajo de ese núcleo central. Cuando la biosfera es considerada el componente principal del cosmos, es natural pensar que los animales y las plantas son, al menos, tan importantes como las rocas y las estrellas en el gran esquema de las cosas, en especial si se sabe muy poco de física o biología. La ciencia moderna ha llegado a una conclusión casi totalmente opuesta. La revolución copernicana convirtió a la Tierra en mero satélite de un Sol central e inanimado. Los posteriores descubrimientos en física y astronomía no sólo demostraron que el universo es vasto en comparación con la Tierra, sino que está perfectamente descrito mediante leyes globales que no hacen alusión a la vida para nada. La teoría de la evolución de Charles Darwin explicó el origen de la vida en términos que no requerían una física especial, y desde entonces son muchos los mecanismos detallados de la vida que se han ido descubriendo, sin que en ninguno de ellos se haya hallado tampoco una física especial.
Esos éxitos espectaculares de la ciencia, así como la gran generalidad de la física newtoniana y, en particular, de la física subsiguiente, contribuyeron en buena medida a hacer atractivo el reduccionismo. Puesto que la fe en la verdad revelada había demostrado ser incompatible con la racionalidad (que requiere apertura a la crítica), muchas personas anhelaban hallar una base definitiva para las cosas en la que pudiesen creer. Si bien aún no disponían de una «teoría total» reductiva en la que creer, no por ello dejaban de aspirar a tenerla. Se dio por sentado que una jerarquía reduccionista de las ciencias, basada en la física subatómica, era inherente a la visión científica del mundo, y, en consecuencia, quienes la criticaban no eran más que pseudocientíficos y seres rebeldes que no querían aceptar la autoridad de la ciencia. Por ello, para cuando yo estudié biología en la escuela, el status de esta materia se había convertido en el contrario del que Aristóteles consideraba evidente. La vida ya no era considerada fundamental. Incluso el término «estudio de la naturaleza» —en el sentido de biología— se había convertido en un anacronismo. Fundamentalmente, la naturaleza era física. Aunque simplifico y exagero un poco, la visión dominante era la que expongo a continuación. La física tenía una ramificación, la química, que estudiaba la interacción de los átomos. La química tenía a su vez una ramificación, la química orgánica, que estudiaba las propiedades de los compuestos del elemento carbono. La química orgánica tenía, asimismo, una ramificación, la biología, que estudiaba los procesos químicos que denominamos vida. Sólo porque resulta que somos uno de esos procesos era interesante para nosotros esa remota ramificación de una materia fundamental. La física, en cambio, se consideraba, evidentemente, importante por derecho propio, ya que el universo todo —incluida la vida— se conforma a sus principios.
Mis condiscípulos y yo debimos aprendernos de memoria una serie de «características de las cosas vivas». Éstas eran meramente descriptivas y hacían escasa referencia a conceptos fundamentales. Por supuesto, la locomoción o movimiento era una de ellas —un mal definido eco de la idea aristotélica—, y tampoco faltaban la respiración y la excreción. Estaban también la reproducción, el crecimiento y una de nombre inolvidable, la irritabilidad, en el sentido de que, «si lo golpeas, devuelve el golpe». Lo que les faltaba a esas supuestas características de la vida en exactitud científica y profundidad no lo compensaba la precisión terminológica. Como había dicho el doctor Johnson, todo objeto real es «irritable». Por otro lado, los virus no respiran, crecen, excretan o se mueven (a menos de ser «golpeados»), y, sin embargo, están bien vivos. Asimismo, los seres humanos estériles no se reproducen, y no por ello están menos vivos.
La razón por la que tanto la concepción de Aristóteles como la de los libros de texto de mi época ni siquiera eran capaces de hacer una buena distinción taxonómica entre cosas vivas y no vivas —y no hablemos de cualquier distinción más profunda—, es que ninguna de las dos se percataba de qué es, en realidad, una entidad viva (un error más perdonable en tiempo de Aristóteles, ya que entonces todo el mundo opinaba como él). La biología moderna no trata de definir la vida mediante algún atributo físico o alguna sustancia —como una «esencia» vital—, características de los que tan sólo la materia viva está dotada. No se espera ya encontrar esencia alguna, puesto que sabemos que la «materia animada» —la materia en forma de organismos vivos— no constituye la base de la vida. Es meramente uno de los efectos de la vida, cuya base es molecular. Es un hecho que existen determinadas moléculas que hacen que determinados entornos las copien.
Estas moléculas reciben el nombre de replicantes. Más generalmente, un replicante es una entidad que hace que ciertos entornos la copien. No todos los replicantes son biológicos, ni todos son moléculas. Un programa informático autocopiante (como un virus informático), por ejemplo, es también un replicante. Un buen chiste es igualmente un replicante, puesto que provoca que los que lo han escuchado lo mencionen para repetirlo ante otras audiencias. Richard Dawkins ha acuñado el término meme para designar los replicantes que son ideas humanas, como los chistes. Pero toda la vida sobre la Tierra se basa en replicantes que son moléculas. Se llaman genes, y la biología es el estudio de sus orígenes, estructura y funcionamiento, así como de sus efectos sobre otra materia. En la mayoría de los organismos, un gen consiste en una secuencia de moléculas más pequeñas, de las que hay cuatro clases distintas, unidas formando una cadena. Los nombres de las moléculas componentes (adenina, citosina, guanina y timina) se utilizan habitualmente abreviados: A, C, G y T. El nombre químico abreviado de una cadena con cualquier número de moléculas A, C, G y T, dispuestas en cualquier orden, es ADN o ácido desoxirribonucleico.
Los genes son verdaderos programas informáticos, escritos como secuencias de los símbolos A, C, G y T, en un lenguaje estándar que se denomina código genético, el cual, con muy ligeras variaciones, es común a toda la vida sobre la Tierra. (Algunos virus se basan en un tipo de molécula relacionada, el ARN o ácido ribonucleico, mientras que los priones son, en cierto sentido, proteínas moleculares autorreplicantes). Determinadas estructuras operan en el interior de las células como ordenadores que ejecutan esos programas genéticos. Dicha ejecución consiste en la obtención de ciertas moléculas (proteínas) a partir de moléculas más simples (aminoácidos), en determinadas condiciones externas. La secuencia «ATG», por ejemplo, es una instrucción para incorporar el aminoácido metionina a la molécula de proteína que se está obteniendo.
De modo característico, un gen es «activado» químicamente en determinadas células del cuerpo, a las que instruye para fabricar la correspondiente proteína. Por ejemplo, la hormona insulina, responsable del control del nivel de azúcar en la sangre de los vertebrados, es una de esas proteínas. Si bien el gen para su fabricación está presente en casi todas las células del cuerpo, sólo es activado en determinadas células especializadas del páncreas, y ello únicamente cuando es necesario. A nivel molecular, eso es todo lo que puede hacer un gen: programar a su ordenador celular para fabricar un determinado compuesto químico. Sin embargo, los genes consiguen actuar como replicantes porque esos programas químicos de bajo nivel se suman, mediante capas y más capas de complejos controles y retroalimentadores, hasta convertirse en complicadísimas instrucciones de alto nivel. Conjuntamente, el gen de la insulina y los genes involucrados en su activación y desactivación constituyen un completo programa para la regulación del nivel de azúcar en el torrente sanguíneo.
Del mismo modo, existen genes que contienen instrucciones específicas que determinan cómo y cuándo deben ser copiados, ellos y otros genes, así como instrucciones para la fabricación de nuevos organismos de la misma especie, incluyendo los ordenadores moleculares que ejecutarán todas esas instrucciones generación tras generación. Hay también instrucciones acerca de cómo debe responder el organismo en su totalidad ante los estímulos, como por ejemplo, cuándo ha de cazar, comer, copular, luchar o huir. Y así sucesivamente.
Un gen puede actuar como replicante tan sólo en determinados entornos. Por analogía con un «nicho» ecológico (el conjunto de entornos en los que un organismo puede sobrevivir y reproducirse), utilizaré también el término nicho para referirme al conjunto de todos los posibles entornos en que un determinado replicante podría hacer que lo copiasen. El nicho de un gen de la insulina incluye entornos en los que está ubicado en el núcleo de una célula en compañía de otros genes, y la célula, por su parte, se encuentra adecuadamente situada en el interior de un organismo funcional, en un hábitat adecuado para su vida y su reproducción. Pero hay también otros entornos capaces de copiar el gen de la insulina, como los laboratorios de biotecnología, en los que bacterias son alteradas genéticamente para incorporarles el gen. Dichos entornos también forman parte del nicho del gen, al igual que otros posibles entornos, infinitamente numerosos, todos ellos muy distintos de aquel en que evolucionó.
No todo lo que puede ser copiado es un replicante. Éste causa que su entorno lo copie, es decir, contribuye causalmente a su propia copia. (Mi terminología difiere ligeramente de la utilizada por Dawkins, quien denomina replicante a todo aquello que es copiado, cualesquiera que sea la razón. Lo que yo denomino replicante, Dawkins lo denominaría replicante activo). Volveré más adelante sobre el significado general de contribuir causalmente a algo, pero lo que quiero decir aquí es que la presencia del replicante y su forma física específica marcan la diferencia entre si va a ser copiado o no. En otras palabras, el replicante es copiado si está presente, pero si fuese reemplazado por otro objeto, incluso muy similar, no lo sería. El gen de la insulina, por ejemplo, provoca solamente un pequeño paso en el enormemente complicado proceso de su propia replicación (ese proceso es el ciclo vital completo del organismo). Pero la inmensa mayoría de las variantes de dicho gen no podrían instruir a las células para elaborar un compuesto químico capaz de realizar las funciones de la insulina. Si los genes de la insulina de un organismo fuesen reemplazados por moléculas ligeramente distintas, dicho organismo moriría (a menos que fuese mantenido en vida por otros medios), y, por consiguiente, no podría reproducirse, de modo que las moléculas en cuestión no podrían ser copiadas. Así pues, que la copia tenga lugar o no es algo extraordinariamente sensible a la forma física del gen de la insulina. Su presencia en la forma y el lugar adecuados marca la diferencia entre si se realizará la copia o no, lo que lo convierte en un replicante, si bien existen innumerables otras causas que contribuirán también a su replicación.
Junto con los genes, en el ADN de la mayoría de los organismos están presentes secuencias aleatorias de A, C, G y T, denominadas a veces secuencias basura del ADN. Son también copiadas y transmitidas a la descendencia. Sin embargo, si semejante secuencia es sustituida por casi cualquier otra de parecida longitud sigue siendo copiada, por lo que podemos deducir que la copia de estas secuencias no depende de su forma física específica. A diferencia de los genes, las secuencias basura no son programas. Si tienen alguna función (y no se sabe aún que la tengan), no puede ser la de transmitir información de ninguna clase. Si bien son copiadas, no contribuyen causalmente a su copia, por lo que no pueden ser consideradas replicantes.
En realidad, esto es una exageración. Todo lo que ha sido copiado debe haber hecho alguna contribución causal a su copia. Las secuencias basura, por ejemplo, están hechas de ADN, lo que permite al ordenador celular copiarlas. No puede copiar moléculas que no sean de ADN. No obstante, no suele resultar ilustrativo considerar replicante a algo que tenga tan sólo una pequeña contribución causal a su propia réplica. Estrictamente hablando, ser replicante es cuestión de grado. Definiré el grado de adaptación de un replicante a un determinado entorno como el grado en el que el replicante contribuye causalmente a su propia réplica en dicho entorno. Cuando un replicante está bien adaptado a la mayoría de los entornos de un nicho, podemos definirlo como bien adaptado al nicho. Hemos visto que el gen de la insulina está altamente adaptado a su nicho. Las secuencias basura tienen un grado despreciable de adaptación en comparación con el del gen de la insulina o cualquier gen fidedigno, pero están mucho más adaptadas a dicho nicho que la mayoría de las demás moléculas.
Téngase en cuenta que, para cuantificar grados de adaptación, debemos tomar en consideración no sólo el replicante en cuestión, sino también el abanico de sus posibles variantes. Cuanto más sensible a la estructura física precisa del replicante sea el proceso de copia en un determinado entorno, más adaptado estará dicho replicante a ese entorno. Para los replicantes altamente adaptados (los únicos que merecen esa denominación) debemos considerar sólo ligeras variaciones, puesto que si se dieran variaciones más amplias ya no serían replicantes. Consideraremos, pues, la sustitución del replicante por objetos más o menos semejantes. Para cuantificar el grado de adaptación de un replicante a un nicho, debemos tener en cuenta su grado de adaptación a todos los entornos del nicho. Deberemos considerar, pues, tanto las variantes del entorno como las del replicante. Si la mayoría de las variantes del replicante no consiguen causar que la mayoría de los entornos las copien, se desprende que la forma del replicante en cuestión es la causa determinante de su propia copia en aquel nicho, que es lo que queremos decir con que está altamente adaptado a él. Por otro lado, si la mayoría de las variantes del replicante son copiadas en la mayoría de los entornos del nicho, ello significa que la forma de ese replicante no incide en la copia, que se producirá en cualquier caso. En consecuencia, el replicante tiene escasa contribución causal a su propia copia y, por lo tanto, no está altamente adaptado a este nicho.
Así pues, el grado de adaptación de un replicante depende no sólo de lo que haga en su entorno concreto, sino también de lo que harían múltiples objetos —la mayoría de los cuales no existen— en un amplio número de entornos distintos del concreto. Ya nos hemos encontrado antes con esta curiosa propiedad. Por ejemplo, la fidelidad de una representación en realidad virtual depende no sólo de las respuestas efectivas de la máquina a las acciones reales del usuario, sino también de las respuestas que no tiene, llegado el caso, a acciones que el usuario no hace de hecho. Esta similitud entre los procesos vitales y la realidad virtual no es pura coincidencia, como explicaré en breve.
El factor más importante que determina el nicho de un gen es, normalmente, que su replicación depende de la presencia de otros genes. La réplica de un gen de insulina de un oso, por ejemplo, depende no sólo de la presencia en el cuerpo del oso de todos sus restantes genes, sino también de la presencia en el medio exterior de otros genes de otros organismos. Los osos no pueden sobrevivir sin comida, y los genes que fabrican esa comida existen solamente en otros organismos.
A menudo, distintas clases de genes que se necesitan mutuamente para su replicación conviven unidos en largas cadenas del ADN: el ADN de un organismo. Un organismo es la clase de cosa —un animal, una planta o un microbio, por ejemplo— que, en términos cotidianos, consideramos viva. Sin embargo, de lo que he dicho anteriormente se desprende que «viva» es, en el mejor de los casos, una manera de hablar cuando la aplicamos a cualquier parte de un organismo que no sea su ADN. Un organismo no es un replicante, sino una parte del entorno de replicantes; habitualmente la parte más importante, después de los demás genes. El resto del entorno lo constituyen la clase de hábitat ocupado por el organismo (por ejemplo, cimas montañosas o profundidades oceánicas) y el estilo de vida particular (caza, microfagia, etcétera) que, dentro de ese hábitat, capacita a dicho organismo para vivir el tiempo suficiente a fin de que sus genes puedan ser replicados.
En el lenguaje cotidiano, decimos que los organismos «se reproducen». No olvidemos que ésta era una de las supuestas «características de las cosas vivas». En otras palabras, pensamos que los organismos son replicantes, pero esto es un error. Los organismos no son copiados durante la reproducción, ni son los causantes de su propia copia. Son formados de nueva planta de acuerdo con planos incorporados al ADN de los organismos de sus progenitores. Por ejemplo, un oso con el hocico deformado por un accidente tal vez cambie de estilo de vida a causa de su deformidad, lo cual podrá afectar para bien o para mal a su capacidad para sobrevivir y «reproducirse». Pero el oso con el hocico deforme no tiene ninguna posibilidad de ser copiado. Si tiene descendencia, ésta tendrá hocicos con la forma original. Sin embargo, si efectuamos alguna modificación en el correspondiente gen (sólo deberemos cambiar una molécula, si lo hacemos inmediatamente después de la concepción) del oso, su descendencia no sólo tendrá hocicos deformes, sino también copias del nuevo gen. Esto demuestra que la nueva forma de los hocicos de esos osos está causada por ese gen, y no por la deformidad de ningún hocico. Así pues, la configuración del hocico del oso progenitor no condiciona la que tendrá el de su descendencia, mientras que la forma de sus genes contribuye, en cambio, a su propia replicación y a la conformación del hocico de sus descendientes.
Por lo tanto, un organismo es el entorno inmediato que copia a los auténticos replicantes: los genes de ese organismo. Tradicionalmente, el hocico de un oso y su madriguera habían sido clasificados respectivamente, como una entidad viva y una entidad no viva, pero esta distinción no está fundada en ninguna diferencia significativa. El papel del hocico del oso no es, en el fondo, distinto del de su madriguera. Ninguna de las dos cosas es un replicante, por más que aparezcan nuevas copias de ambas de modo constante. Tanto el hocico como la madriguera son meras partes del entorno que manipulan los genes del oso durante el proceso de hacerse replicar.
La noción de que la vida se basa en los genes —que considera a los organismos como parte del entorno de los genes— ha constituido, implícitamente, la base de la biología desde Darwin, pero no se le dio la debida importancia, al menos hasta los años sesenta, y no puede decirse que se comprendieran todas sus implicaciones hasta que Richard Dawkins publicó El gen egoísta (1976) y El fenotipo expandido (1982).
Vuelvo a la pregunta de si la vida es o no un fenómeno fundamental de la naturaleza. Ya he advertido contra la asunción reduccionista de que los fenómenos emergentes, como la vida, son necesariamente menos fundamentales que los físicos microscópicos. Sin embargo, todo lo que acabo de decir sobre la vida parece indicar que es un mero efecto secundario más, al final de una larga cadena de efectos secundarios. Y es que no son tan sólo las predicciones de la biología las que se sujetan, en principio, a las de la física, sino también sus explicaciones. Como he dicho, las grandes teorías explicativas de Darwin (en sus versiones modernas, como la de Dawkins) y de la bioquímica moderna son reductivas. Las moléculas vivas —genes— son, después de todo, moléculas, tan sujetas a las leyes de la física y la química como las no vivas. No contienen ninguna sustancia especial, ni presentan ningún atributo físico extraordinario. Simplemente, son, en determinados entornos, replicantes. La propiedad de ser replicante es altamente contextual, es decir, depende de intrincados detalles del entorno. Una entidad puede ser replicante en un entorno y no en otro. Del mismo modo, la propiedad de estar adaptado a un nicho no depende de un simple e intrínseco atributo físico del que pueda hacer uso el replicante en un momento determinado, sino de los efectos que ese nicho pueda causar en él en el futuro en circunstancias hipotéticas (por ejemplo, variantes del entorno). Las propiedades contextuales e hipotéticas son, en esencia, derivativas, de modo que resulta difícil creer que un fenómeno caracterizado únicamente por ellas pueda ser fundamental en la naturaleza.
Por lo que se refiere al impacto físico de la vida, la conclusión es la misma: los efectos de la vida parecen despreciablemente pequeños. Por lo que sabemos, el planeta Tierra es el único lugar del universo en el que existe. El hecho es que no hemos encontrado pruebas de su existencia en ninguna otra parte, de modo que, aun en el supuesto de que estuviese ampliamente extendida, sus efectos serían demasiado pequeños para ser perceptibles para nosotros. Lo que vemos más allá de la Tierra es un universo activo, lleno de procesos diversos y poderosos, pero completamente inanimados. Las galaxias evolucionan. Las estrellas se condensan, brillan, flamean, estallan y se contraen. Partículas de alta energía y ondas electromagnéticas y gravitatorias fluyen en todas direcciones. Que la vida esté presente o no en esos titánicos procesos no parece tener mayor importancia. Todo indica que ninguno de ellos se vería afectado en lo más mínimo si estuviera presente. Si la Tierra se viese envuelta en una llamarada solar —lo que en sí mismo constituiría un insignificante acontecimiento astrofísico—, nuestra biosfera quedaría instantáneamente esterilizada. Esa catástrofe, sin embargo, tendría sobre el Sol el mismo efecto que una gota de lluvia sobre un volcán en erupción. Nuestra biosfera es, en términos de su masa, energía o cualquier otra dimensión astrofísica significativa, algo insignificante, incluso en comparación con la Tierra. Es, además, un tópico en astrofísica que el sistema solar consiste esencialmente en el Sol y Júpiter. Todo lo demás (incluyendo la Tierra) es considerado «impurezas». Más aún, el sistema solar es una porción nimia de nuestra galaxia, la Vía Láctea, inapreciable a su vez entre la multitud de galaxias conocidas en el universo. Parece, pues, que, como dice Stephen Hawking: «La raza humana no es más que escoria química sobre un planeta de dimensiones discretas, el cual órbita alrededor de una estrella nada notable en el borde de una galaxia que no se distingue en absoluto de los cientos de miles de millones de galaxias restantes».
La idea dominante en la actualidad es, pues, que la vida, lejos de ser el centro, ya sea desde el punto de vista geométrico, teorético o práctico, es de una insignificancia casi inconcebible. La biología es, en este contexto, una materia con un status parecido al de la geografía. Conocer la trama urbana de la ciudad de Oxford es importante para los que vivimos en ella, pero carece de todo interés para quienes nunca la visiten. De modo semejante, parece que la vida es una propiedad de una pequeña área, o quizás áreas, del universo, fundamental sólo para nosotros porque estamos vivos, pero sin ninguna trascendencia, teorética o práctica, en el vasto esquema de las cosas.
Pero, por sorprendente que parezca, esta apariencia es engañosa. Es una falsedad que los efectos físicos de la vida sean insignificantes, o que sea un mero derivativo desde el punto de vista teorético.
Como primer paso para explicarlo, permítaseme profundizar en mi anterior afirmación de que la vida es una forma de generación de realidad virtual. He empleado la palabra «ordenadores» para describir los mecanismos que ejecutan los programas de los genes en el interior de las células, pero esta terminología es, en realidad, un tanto ambigua. En comparación con los ordenadores de uso general que fabricamos artificialmente, hacen más en algunos aspectos y menos en otros. No resultaría fácil programarlos para procesar textos o factorizar grandes números. En cambio, ejercen un control interactivo extremadamente exacto sobre las respuestas de un complejo entorno (el organismo) ante todo lo que le pueda suceder. Este control, además, está dirigido a hacer que el entorno realice una retroacción sobre los genes de un modo específico (es decir, replicándolos), de manera que el efecto neto sobre los genes resulte tan independiente como sea posible de lo que esté sucediendo en el exterior. Esto es más que informática, es generación de realidad virtual.
La analogía con la tecnología humana de realidad virtual no es, sin embargo, perfecta. En primer lugar, si bien los genes se hallan inmersos, al igual que el usuario de la realidad virtual, en un entorno cuya constitución y comportamiento detallados están especificados por un programa (al que los propios genes dan forma material), los genes no experimentan ese entorno, puesto que carecen de sentidos y experiencias. En consecuencia, si bien un organismo es una representación en realidad virtual especificada por sus genes, es una representación sin audiencia. En segundo lugar, el organismo no sólo está siendo representado, sino también fabricado. No se trata de «engañar» al gen para hacerle creer que hay un organismo, sino que éste se encuentra realmente allí.
No obstante, esas diferencias son insustanciales. Como he dicho, toda representación en realidad virtual fabrica físicamente el entorno representado. El interior de cualquier generador de realidad virtual en el acto de representar es, precisamente, un entorno físico y real, fabricado para tener las propiedades especificadas en el programa. Somos los usuarios quienes escogemos, en ocasiones, interpretarlo como un entorno distinto, que resulta que provoca las mismas sensaciones. En cuanto a la ausencia de usuario, consideremos explícitamente en qué consiste el papel del usuario de realidad virtual. En primer lugar, debe «golpear» al entorno para que éste lo «golpee» a su vez; en otras palabras, ha de interactuar con el entorno de un modo autónomo. En el caso biológico, ese papel es asumido por el hábitat exterior. En segundo lugar, el usuario debe proporcionar la intención que hay tras la representación. Es decir, no tiene sentido afirmar que una determinada situación constituye una representación en realidad virtual sin tener en cuenta el concepto de fidelidad en la representación. Como dije, la fidelidad de un entorno es la semejanza, percibida por el usuario, entre el entorno reproducido y el que se deseaba reproducir. Pero ¿qué significa la fidelidad si se trata de un entorno que nadie deseaba reproducir y nadie percibe? Significa el grado de adaptación de los genes a su nicho. Podemos inferir la «intención» de los genes de reproducir un entorno que los replique de acuerdo con la teoría de la evolución de Darwin. Los genes se extinguen si no plasman esa «intención» tan eficiente y resueltamente como otros genes que compiten con ellos.
Así pues, los procesos vitales y las representaciones en realidad virtual son, diferencias superficiales aparte, la misma clase de proceso. Ambos implican la incorporación física de teorías generales sobre un entorno. En ambos casos esas teorías son utilizadas para construir el entorno y controlar, de modo interactivo, no tan sólo su apariencia instantánea, sino su respuesta detallada ante estímulos generales.
Los genes dan forma material al conocimiento sobre sus nichos. Todo lo que tiene importancia fundamental en el fenómeno de la vida depende de esa propiedad, y no de la replicación per se. Podemos, pues, llevar ahora la discusión más allá de los replicantes. En principio, podríamos imaginar una especie cuyos genes fuesen incapaces de replicarse, pero que, en cambio, estuviesen adaptados para mantener invariable su aspecto físico mediante automantenimiento continuo y protección ante las influencias externas. No es probable que una especie así evolucionase de manera espontánea, pero podría ser obtenida de modo artificial. Al igual que el grado de adaptación de un replicante se define como el grado en que contribuye causalmente a su propia replicación, podemos definir el grado de adaptación de esos genes no replicantes como el grado en que contribuyen a su propia supervivencia en una determinada forma. Imaginemos una especie cuyos genes fuesen estructuras grabadas en la superficie de un diamante en bruto. Un diamante en bruto, tenga la forma que tenga, puede sobrevivir casi eternamente en gran variedad de circunstancias, pero esa forma no está adaptada para la supervivencia, ya que un diamante con una forma distinta sobreviviría también en circunstancias similares. Pero si los genes codificados en un diamante de nuestra especie hipotética fueran los causantes de que ese organismo se comportase de una manera determinada que, por ejemplo, protegiese la superficie grabada del diamante de la corrosión de un entorno hostil, o la defendiese de otros organismos que tratasen de introducir información distinta, o de ladrones que lo quisieran tallar y pulir para convertirlo en una gema, contendría entonces genuinas adaptaciones para la supervivencia en dichos entornos. (Incidentalmente, una gema posee cierto grado de adaptación para la supervivencia en el actual entorno terrestre. Los humanos buscan los diamantes en bruto y transforman sus formas naturales en gemas, pero también buscan las gemas y respetan sus formas. Así pues, en ese entorno, la forma de una gema contribuye causalmente a su supervivencia).
Si cesase la fabricación de esos organismos artificiales, no podría volver a incrementarse el número de casos de cada gen no replicante, pero tampoco decrecería mientras el conocimiento que contuviesen fuera suficiente para permitirles mantener su estrategia de supervivencia para el nicho ocupado. Con el tiempo, un cambio lo bastante importante en el hábitat, o el deterioro causado por accidentes, podría conllevar la eliminación de la especie, pero, en caso contrario, ésta podría muy bien sobrevivir tanto como cualquier especie natural. Los genes de esas especies comparten las propiedades de los genes reales, excepto la replicación. En particular, incorporan el conocimiento necesario para «simular» sus organismos, es decir, reproducirlos, del mismo modo en que lo hacen los genes reales.
Es la supervivencia del conocimiento, y no por fuerza la de los genes o cualquier otro objeto físico, lo que constituye el denominador común entre genes replicantes y no replicantes. Estrictamente hablando, pues, lo que está o no adaptado a cierto nicho es, en realidad, un fragmento de conocimiento, más que un objeto físico. Si está adaptado, tendrá la propiedad de que, una vez establecido en ese nicho, tenderá a permanecer allí. En el caso de un replicador, el material físico que le da forma cambia sin cesar, y con cada replicación se forma una nueva copia compuesta de partes no replicantes. El conocimiento no replicante puede también materializarse sucesivamente en distintas formas físicas, como, por ejemplo, cuando se transfiere el sonido de un disco antiguo, primero a una cinta de audio y luego a un disco compacto. Podríamos imaginar otros organismos vivos artificiales basados en no replicantes que se comportasen del mismo modo, aprovechando cada oportunidad de recopiar el conocimiento de sus genes para incorporarlo al medio más seguro disponible. Quizás algún día nuestros descendientes lo hagan.
No creo que resulte justo denominar «inanimados» a estos organismos, pero la terminología no es lo más importante, sino el hecho de que, si bien la vida toda se basa en replicantes, de lo que realmente trata el fenómeno de la vida es del conocimiento. Podemos, pues, dar una definición de adaptación directamente en términos de conocimiento: Una entidad está adaptada a su nicho cuando da forma material al conocimiento que causa que ese nicho mantenga la existencia de dicho conocimiento. Nos estamos acercando a la razón por la que la vida es fundamental. La vida trata de la materialización física del conocimiento. En el capítulo 6 nos encontramos con una ley de física, el principio de Turing, que trata también de la materialización física del conocimiento. Nos dice que es posible dar forma material a las leyes de la física, en cuanto son de aplicación a cualquier entorno físicamente posible, a los programas de un generador de realidad virtual. Los genes son programas así. Aún más, cualesquiera otros programas de realidad virtual que existan, o puedan llegar a existir físicamente, son efectos directos o indirectos de la vida. Los programas de realidad virtual que ejecutan nuestros ordenadores o nuestros cerebros, por ejemplo, son efectos indirectos de la vida humana. La vida es, pues, el medio, presumiblemente necesario, mediante el cual los efectos a los que se refiere el principio de Turing son aplicados en la naturaleza.
Si bien ello resulta alentador, no es suficiente para establecer que la vida sea un fenómeno fundamental. Eso se debe a que no he demostrado aún que el principio de Turing tenga el status de ley fundamental. Un escéptico podría argumentar que no lo tiene. Se trata de una ley sobre la materialización física del conocimiento, y el escéptico podría adoptar la postura de que el conocimiento no es algo fundamental, sino un limitado concepto antropocéntrico. En otras palabras, se trata de una de esas cosas importantes para nosotros por lo que somos —animales cuyo nicho ecológico depende de la creación y aplicación de conocimiento—, pero no significativas en un sentido absoluto. Para un oso koala, cuyo nicho ecológico depende de las hojas del eucalipto, dicho árbol es significativo. Para el simio Homo sapiens, manipulador de conocimiento, éste es significativo.
Pero el escéptico estaría en un error. El conocimiento no es significativo únicamente para el Homo sapiens, ni sólo en el planeta Tierra. Como he dicho, el que algo tenga o no un gran impacto físico no es determinante, pero sí relevante, para que sea importante en la naturaleza. Consideremos ahora los efectos astrofísicos del conocimiento.
La teoría de la evolución estelar —la estructura y el desarrollo de las estrellas— es uno de los grandes éxitos de la ciencia. (Fíjense en el conflicto terminológico: la palabra «evolución» significa en física desarrollo o, simplemente, movimiento, no variación ni selección). Hace nada más un siglo, se desconocía incluso el origen de la energía solar. La física más avanzada de la época sólo proporcionaba la errónea conclusión de que, cualquiera que fuese su fuente de energía, el Sol no podía haber estado brillando durante más de cien millones de años. Resulta curioso que geólogos y paleontólogos ya sabían, por la evidencia fósil del desarrollo de la vida, que el Sol debía de haber estado brillando sobre la Tierra durante mil millones de años, por lo menos. Luego se descubrió la física nuclear y fue aplicada con gran detalle a la física del interior de las estrellas. Desde entonces, la teoría de la evolución estelar ha ido madurando. Comprendemos ahora lo que provoca el brillo de las estrellas. Para la mayoría de clases de estrellas podemos predecir qué temperatura, color, luminosidad y diámetro presentarán en cada etapa de su historia, cuánto durarán, qué elementos formará la estrella mediante transmutaciones nucleares, etcétera. Esta teoría ha sido comprobada y confirmada mediante observaciones del Sol y otras estrellas.
Podemos, pues, utilizar esta teoría para predecir el futuro desarrollo del Sol. Nos dice que el Sol continuará brillando con gran estabilidad durante otros cinco mil millones de años, más o menos, luego se expandirá hasta alcanzar un diámetro un centenar de veces superior al actual y se transformará en una estrella gigante roja; luego empezará a centellear, su brillo aumentará repentinamente, con lo que pasará a ser una nova, se contraerá y se enfriará, hasta convertirse con el tiempo en una enana negra. Pero ¿le ocurrirá de veras todo esto al Sol? ¿Se han convertido ya todas las estrellas formadas unos cuantos miles de millones de años antes que el Sol, con idéntica masa y composición, en gigantes rojas, como predice la teoría? ¿Es posible que algunos procesos químicos en apariencia insignificantes, sucedidos en los planetas menores en órbita alrededor de esas estrellas, puedan haber alterado el curso de procesos nucleares y gravitatorios de masa y energía infinitamente superiores?
Si el Sol se convierte en una gigante roja, absorberá a la Tierra y la destruirá. Si algunos de nuestros descendientes, físicos o intelectuales, se encuentran aún sobre la Tierra por aquel entonces, quizás no deseen que eso ocurra. Harán, sin duda, todo lo que esté en su mano para impedirlo.
¿Es incuestionablemente cierto que no podrán hacer nada al respecto? Sin duda, nuestra tecnología actual es demasiado insignificante para ello, pero ni nuestra teoría de la evolución estelar, ni ninguna otra física conocida, nos da motivo para pensar que la tarea sea imposible. Bien al contrario, sabemos ya, en términos amplios, lo que ésta conllevaría (básicamente, extraer materia del Sol). Nos quedan varios miles de millones de años para perfeccionar nuestros planes ahora en ciernes y ponerlos en práctica. Si, llegado el caso, nuestros descendientes consiguieran salvarse de ese modo, ello significaría que nuestra teoría actual de la evolución estelar, aplicada en concreto a la estrella Sol, hace una predicción completamente equivocada. Y la razón para ello es que no toma en consideración el efecto de la vida en la evolución estelar. Tiene en cuenta efectos físicos fundamentales, como las fuerzas nucleares y electromagnéticas, la gravedad, la presión hidrostática y la de la radiación, pero no la vida.
Parece razonable pensar que el conocimiento necesario para modificar el Sol de ese modo no puede evolucionar únicamente por selección natural, de modo que cualquier futura modificación del Sol tiene que depender de la vida inteligente. Ante esto, se puede argumentar que quizás sea una desmesurada e infundada asunción que la inteligencia pueda sobrevivir sobre la Tierra durante varios miles de millones de años y que, incluso en este caso, es también una asunción pensar que se posea entonces el conocimiento necesario para modificar el Sol. Una opinión corriente en la actualidad es que la vida inteligente sobre la Tierra corre el peligro de autodestruirse, bien como consecuencia de una guerra nuclear, bien por algún efecto secundario del avance tecnológico o la investigación científica. Muchas personas creen que si la vida inteligente sobrevive, sólo lo conseguirá suprimiendo el progreso tecnológico. Podrían, pues, temer que el desarrollo de la tecnología necesaria para la modificación de las estrellas fuera incompatible con la supervivencia durante el tiempo suficiente para utilizarla, y que, por consiguiente, la vida sobre la Tierra está predestinada, de un modo u otro, a no afectar a la evolución del Sol.
Estoy convencido de que semejante pesimismo es infundado y, como explicaré en el capítulo 14, tenemos todos los motivos para conjeturar que, con el tiempo, nuestros descendientes podrán modificar el Sol y hacer muchas más cosas. Sin duda, no podemos prever ni su tecnología ni sus deseos. Puede que escojan salvarse emigrando del sistema solar, o refrigerando la Tierra, o utilizando cualquier otro medio, inconcebible hoy para nosotros, qué no requiera intervenir en el Sol. Por otra parte, podrían decidir modificarlo mucho antes de lo necesario, para prevenir su entrada en la fase de gigante roja (dominando su energía más eficazmente, por ejemplo, o explotando para mejorar su calidad de vida los minerales que contiene en su interior). Sin embargo, lo que quiero dejar bien claro es que lo que ocurra no dependerá de nuestra capacidad de predicción, sino sólo de qué conocimiento tengan nuestros descendientes y cómo decidan aplicarlo. No podemos, pues, predecir el futuro del Sol sin tomar posición sobre el futuro de la vida sobre la Tierra, en general, y del conocimiento, en particular. El color que tenga el Sol dentro de diez mil millones de años depende de la presión gravitatoria y radiactiva, la convección y la síntesis nuclear. No depende en absoluto de la geología de Venus, la química de Júpiter o la disposición de los cráteres de la Luna. Pero depende de lo que le ocurra a la vida inteligente en el planeta Tierra. Depende de la política, la economía y el resultado de las guerras. Depende de lo que haga la gente, de qué decisiones tome, qué problemas resuelva, qué valores tenga y cómo enfoque el bienestar futuro de sus hijos.
No podemos evitar llegar a esta conclusión adoptando una teoría pesimista sobre las perspectivas de nuestra supervivencia. Semejante teoría no se desprende de las leyes de la física ni de ningún otro principio fundamental que conozcamos, y tan sólo puede ser justificada en términos humanos de alto nivel, tales como que «el conocimiento científico ha sobrepasado al conocimiento moral», o afirmaciones por el estilo. Al utilizar esa teoría como argumentación se reconoce de un modo implícito que las teorías sobre los asuntos humanos son necesarias para efectuar predicciones astrofísicas. Incluso en el supuesto de que la especie humana fracasase en sus esfuerzos por sobrevivir, ¿sería de aplicación la teoría pesimista a toda clase de inteligencia extraterrestre del universo? En caso contrario, si alguna clase de vida inteligente, en alguna galaxia, llegara a sobrevivir durante miles de millones de años, confirmaría que la vida es significativa en el desarrollo físico general del universo.
A lo largo y lo ancho de nuestra galaxia y el multiverso, la evolución estelar depende de si se ha desarrollado la vida inteligente y, en caso afirmativo, de dónde lo ha hecho, de los resultados de sus guerras y de cómo enfoca el futuro de sus hijos. Podemos, por ejemplo, predecir aproximadamente las proporciones de estrellas de distintos colores (o, más precisamente, de distintos tipos espectrales) que debería haber en nuestra galaxia. Para ello, tendremos que hacer algunas asunciones sobre cuánta vida inteligente hay allí y cuáles son sus manifestaciones (¡saber cuántas estrellas ha sido capaz de apagar ayudaría mucho!). Por ahora, nuestras observaciones dan como resultado que no existe vida inteligente fuera del sistema solar. Cuando nuestras teorías sobre la estructura de la galaxia de la que formamos parte estén más depuradas, podremos hacer predicciones más exactas, pero, de nuevo, únicamente sobre la base de asunciones sobre la distribución y el comportamiento de la inteligencia en ella. Si estas asunciones son incorrectas, haremos predicciones erróneas sobre la distribución de los tipos espectrales. Pero, si son correctas y detectamos ciertas anomalías en la distribución de dichos tipos espectrales, ello podría ser una prueba de la presencia de inteligencia extraterrestre.
Los cosmólogos John Barrow y Frank Tipler han considerado los efectos astrofísicos que tendría la vida en caso de sobrevivir durante un largo período después que el Sol se hubiese convertido en una gigante roja. Han descubierto que, con el tiempo, causaría importantes cambios cualitativos en la estructura de nuestra galaxia y, más tarde, en la de todo el universo (volveré sobre estos resultados en el capítulo 14). Una vez más, cualquier teoría sobre la estructura del universo, en lo que no sean sus etapas iniciales, deberá tomar posición sobre lo que la vida hará o dejará de hacer en cada momento. No hay escapatoria posible: la historia futura del universo depende de la historia futura del conocimiento. Los astrólogos creían que los acontecimientos cósmicos influían en los asuntos humanos, mientras que la ciencia ha creído durante siglos que no se afectaban mutuamente. Ahora vemos que son los asuntos humanos los que influyen en los acontecimientos cósmicos.
Vale la pena reflexionar qué nos hizo subestimar el impacto físico de la vida. Fue el tener una visión demasiado restringida de las cosas, lo que no deja de ser irónico, puesto que los antiguos llegaron a un consenso que conseguía evitar nuestro error precisamente por tener una visión de las cosas aún más restringida. En el universo, tal como lo vemos, la vida no ha tenido ningún efecto astrofísico importante. Sin embargo, únicamente vemos el pasado, y, dentro de él, sólo con cierto detalle lo que se halla más próximo a nosotros espacialmente. Cuanto más lejos miramos en el universo, más nos adentramos en el pasado y menos detalle percibimos. Pero el pasado, incluso considerado en su totalidad —la historia del universo desde el Big Bang hasta nuestros días—, es tan sólo una mínima parte de la realidad física. Queda por transcurrir un espacio de tiempo muchísimo más largo entre el momento actual y el Big Crunch (si llega a suceder), por no mencionar los restantes universos. No podemos observarlo, pero cuando aplicamos nuestras mejores teorías al futuro de las estrellas, las galaxias y el universo, percibimos un amplio campo en el que la vida puede influir y, a largo plazo, llegar a dominar todo lo que ocurra, al igual que hace ahora en la biosfera de la Tierra.
El argumento convencional para justificar la insignificancia de la vida otorga demasiado peso a las cantidades observables y «voluminosas» como la dimensión, la masa y la energía. Para la visión restringida pasada y presente, ésas eran, y siguen siéndolo, buenas medidas de la importancia astrofísica, pero no existe razón alguna en el campo de la física para que continúe siendo así. Es más, la propia biosfera proporciona abundantes ejemplos contrarios a la aplicación general de dichas medidas de importancia. En el siglo III antes de Cristo, por ejemplo, la masa de la población humana era de unos diez millones de toneladas. Podríamos pensar que no es probable que los procesos físicos ocurridos en dicho siglo, que involucraban el movimiento de masas muchas veces superiores, pudiesen verse significativamente afectados por la presencia o la ausencia de seres humanos. Sin embargo, la gran muralla china, cuya masa es de unos trescientos millones de toneladas, fue construida en esa época. Mover millones de toneladas de rocas es una acción que los humanos realizan constantemente. En la actualidad, sólo son necesarias algunas docenas de humanos para remover una masa de un millón de toneladas en la explanación de una vía férrea o la perforación de un túnel. Ello se pone aún más de relieve si establecemos la comparación, todavía más lógica, entre la masa de la tierra y las rocas desplazadas y la de la pequeña parte del cerebro del emperador o del ingeniero de la que surgen las ideas (o memes) que harán que dicha masa sea removida. La especie humana, entendida como un todo (o, si se prefiere, su reserva de memes), dispone ya, probablemente, del conocimiento suficiente para destruir varios planetas, si su supervivencia dependiese de ello. Incluso la vida no inteligente ha transformado la superficie y la atmósfera de la Tierra de un modo que equivale a muchas veces su propia masa. Todo el oxígeno de nuestra atmósfera, por ejemplo, ha sido creado por las plantas y es, por consiguiente, un efecto secundario de la replicación de los genes (es decir, de las moléculas), que descendían de una sola molécula. La vida no obra sus efectos por ser mayor o tener más masa o más energía que otros procesos físicos, sino por poseer más conocimiento. En términos de efecto final sobre los resultados de los procesos físicos, el conocimiento es, al menos, tan importante como cualquier otra cantidad física.
Ahora bien: ¿existe en el caso de la vida, como asumían en la antigüedad que debía ocurrir, una diferencia física básica entre los objetos portadores de conocimiento y los no portadores, una diferencia que no depende ni de los entornos del objeto ni de sus efectos en el futuro remoto, sino únicamente de sus atributos físicos inmediatos? Por extraordinario que parezca, la hay. Para ver en qué consiste, debemos adoptar la perspectiva del multiverso.
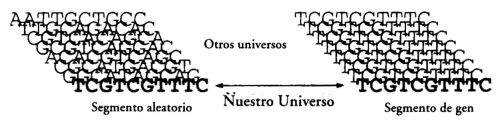
Consideremos el ADN de un organismo vivo, por ejemplo, un oso, y supongamos que en alguna parte de uno de sus genes encontramos la secuencia TCGTCGTTTC. Esta cadena concreta de diez moléculas, tanto en el nicho especial formado por el resto del gen como en su propio nicho, es un replicante. Incorpora una pequeña, pero importante, cantidad de conocimiento. Supongamos ahora, para redondear la hipótesis, que encontramos un segmento de ADN basura (de no gen) en el ADN del oso que tiene también la secuencia TCGTCGTTTC. Esta secuencia, sin embargo, no merece ser considerada replicante, puesto que apenas contribuye a su replicación y no es portadora de conocimiento alguno. Se trata de una secuencia aleatoria. Tenemos, pues, dos objetos físicos, segmentos ambos de la misma cadena de ADN, uno de los cuales es portador de conocimiento, mientras que el otro es una secuencia aleatoria. Sin embargo, son físicamente idénticos. ¿Cómo puede el conocimiento ser una cantidad física fundamental si un objeto lo posee y otro que es idéntico a él no?
Puede serlo porque esos dos segmentos no son, en realidad, idénticos. Lo parecen tan sólo cuando son vistos desde algunos universos, como el nuestro. Veámoslos de nuevo, pero esta vez desde otros universos. Como no lo podemos hacer físicamente, lo haremos desde un punto de vista teórico.
Sabemos que el ADN de los organismos vivos está sujeto naturalmente a variaciones aleatorias —mutaciones— en la secuencia de las moléculas A, C, G y T. Según la teoría de la evolución, en los genes las adaptaciones y, por consiguiente, su propia subsistencia, dependen de que dichas mutaciones hayan tenido lugar. Gracias a ellas, las poblaciones de cualquier gen contienen un determinado grado de variaciones, y los individuos portadores de genes con niveles más altos de adaptación son los que tienden a tener más descendencia. La mayoría de las variaciones de un gen lo incapacitan para su replicación, puesto que la secuencia alterada ya no transmite las instrucciones a la célula para producir algo útil. Otras variaciones, simplemente, hacen menos probable la replicación, es decir, disminuyen el nicho del gen.
Algunas, sin embargo, pueden ser portadoras de nuevas instrucciones que hagan más probable la replicación. De este modo tiene lugar la selección natural. Con cada generación de variaciones y replicaciones el grado de adaptación de los genes supervivientes tiende a incrementarse. Ahora bien, una mutación aleatoria, causada, por ejemplo, por el impacto de un rayo cósmico, origina variaciones no sólo dentro de la población del organismo en un universo, sino también entre universos. Un «rayo» cósmico es una partícula subatómica de alta energía que, como el fotón emitido por la linterna, viaja en diferentes direcciones en universos distintos. Así pues, cuando un rayo cósmico impacta en una cadena de ADN y produce una mutación, algunas de sus contrapartidas en otros universos se quedarán sin sus correspondientes copias de esa cadena de ADN, mientras que otras impactarán en distintas posiciones y originarán mutaciones diferentes. De este modo, un solo rayo cósmico que incidiera sobre una sola molécula de ADN causaría, en general, la aparición de gran número de mutaciones diferentes en distintos universos.
Cuando consideramos la posible apariencia de un determinado objeto físico en otros universos, no debemos mirar tan lejos en el multiverso que nos resulte imposible identificar una contrapartida suya en el otro universo. Tomemos, por ejemplo, un segmento de ADN. En algunos universos, simplemente, no hay moléculas de ADN. En otros, existe el ADN, pero son universos tan distintos del nuestro, que no hay modo de identificar qué segmento del ADN que se encuentra en ellos corresponde al que estamos considerando en éste. Resulta ocioso preguntarse qué aspecto tendría nuestro segmento de ADN en semejantes universos, de modo que debemos considerar únicamente universos lo bastante similares al nuestro para que no surja esa disparidad. Podríamos considerar, por ejemplo, tan sólo aquellos universos en los que existan osos y en los que, además, se haya introducido una muestra del ADN de un oso en una máquina de análisis programada para imprimir diez letras que representen su estructura en una posición específica, relativa a ciertas referencias sobre una determinada cadena de ADN. La argumentación que sigue no se vería afectada aunque escogiésemos cualquier otro criterio razonable para identificar segmentos de ADN de características similares en universos cercanos.
Según cualquiera de esos criterios, el segmento del gen del oso debe tener la misma secuencia en casi todos los universos próximos que en el nuestro. Ello se debe a que cabe presumir que está altamente adaptado, lo que significa que la mayor parte de sus variantes no conseguirían hacerse copiar en la mayor parte de las variantes de sus entornos, y no podrían, por tanto, aparecer en dicha localización en el ADN de un oso vivo. En cambio, cuando el segmento de ADN no portador de conocimiento experimenta prácticamente cualquier mutación, la versión mutante seguirá siendo capaz de ser copiada. A lo largo de generaciones de replicaciones habrán ocurrido múltiples mutaciones, la mayoría de las cuales no habrán afectado a la replicación. Por consiguiente, el segmento de ADN basura, a diferencia de su contrapartida en el gen, será completamente heterogéneo en distintos universos. Podría muy bien suceder que cada posible variación de su secuencia estuviera representada por un igual en el multiverso (que es lo que debería entenderse cuando se dice que su secuencia es estrictamente aleatoria).
Por consiguiente, la perspectiva del multiverso revela una estructura física adicional en el ADN del oso. En nuestro universo, contiene dos segmentos con la secuencia TCGTCGTTTC. Uno de ellos es parte de un gen, mientras que el otro no. En casi todos los universos cercanos, el primer segmento conserva la secuencia TCGTCGTTTC que presenta en el nuestro, pero el segundo varía enormemente de secuencia de un universo próximo a otro. Así pues, y desde la perspectiva del multiverso, los dos segmentos no son ni remotamente parecidos (figura 8.1).
FIGURA 8.1. Vista desde el multiverso de dos segmentos de ADN que resultan ser idénticos en nuestro universo, uno aleatorio y el otro del interior de un gen.
Una vez más, tuvimos un punto de vista demasiado restrictivo y nos dejamos llevar a la falsa conclusión de que las entidades portadoras de conocimiento pueden ser físicamente idénticas a las no portadoras, lo cual, a su vez, proyectó dudas sobre el status fundamental del conocimiento. Pero ahora hemos vuelto prácticamente al punto de partida. Reconocemos que la antigua idea de que la materia viva tiene propiedades especiales se aproximaba mucho a la verdad: no es la materia viva, sino la materia portadora de conocimiento, la que es físicamente especial. Dentro de un universo parece irregular, pero en el multiverso presenta una estructura tan regular como la de un cristal.
En definitiva, el conocimiento es una cantidad física fundamental, y el fenómeno de la vida también, aunque en medida ligeramente inferior.
Imaginemos que observamos una molécula del ADN de una célula de oso con un microscopio electrónico y tratamos de distinguir las secuencias de gen de las de no gen, así como de estimar el grado de adaptación de cada gen. En cualquier universo, considerado aisladamente, esa tarea resultaría imposible. La propiedad de ser un gen —es decir, de estar altamente adaptado— es abrumadoramente complicada para que sea posible detectarla en un universo aislado. Se trata de una propiedad emergente. Deberíamos hacer múltiples copias del ADN con sus variaciones, utilizar la ingeniería genética para obtener numerosos embriones de oso para cada variante del ADN, esperar a que los osos crecieran y vivieran en diversos entornos representativas del nicho ecológico de su especie y ver cuáles de ellos conseguían procrear.
Pero con un microscopio mágico, capaz de ver en otros universos (lo que, insisto, no es posible: utilizamos una teoría para imaginar —o representar— lo que sabemos que debe existir allí), la tarea resultaría sencilla. Como en la figura 8.1, los genes se distinguirían de los no genes, del mismo modo que los campos cultivados se distinguen de los bosques en una fotografía aérea, o los cristales que han precipitado se destacan en una solución. Serían regulares en múltiples universos próximos, mientras que los no genes, los segmentos de ADN basura, serían irregulares. En cuanto al grado de adaptación de un gen, éste resultaría casi tan fácil de determinar. Los genes mejor adaptados presentarían la misma estructura en un amplio abanico de universos: tendrían «cristales» más grandes, por así decirlo.
Viajemos ahora a otro planeta para tratar de encontrar las formas de vida autóctonas, si existen. Una vez más, ésta es una tarea de notoria dificultad. Deberíamos realizar complejos y sutiles experimentos, cuyos innumerables fracasos han sido tema de numerosos relatos de ciencia ficción. Pero si pudiéramos, simplemente, observar el planeta con un telescopio «multiversal», la vida y sus consecuencias serían evidentes al primer vistazo. Sólo habría que buscar estructuras complejas que pareciesen irregulares en cualquier otro universo, pero idénticas en muchos universos próximos. Si encontrásemos alguna, habríamos hallado la evidencia de un conocimiento materializado físicamente. Donde hay conocimiento, debe haber vida, al menos en el pasado.
Comparemos ahora a un oso con la constelación de la Osa Mayor. El oso es, anatómicamente, muy similar en muchos universos próximos. No son tan sólo sus genes los que presentan esa propiedad, sino todo su cuerpo (si bien otros atributos de este último, como el peso, variarán mucho más que sus genes; ello se debe a que, por ejemplo, en los distintos universos el oso habrá tenido diferente fortuna en su búsqueda de alimento). Pero en el caso de la constelación de la Osa Mayor no se da esta regularidad de un universo a otro. La configuración de esa constelación es el resultado de las condiciones iniciales del gas galáctico del que se formaron las estrellas. Estas condiciones fueron aleatorias —muy diversas en distintos universos, a nivel microscópico— y el proceso de formación de las estrellas a partir del gas tuvo como consecuencia diversas inestabilidades, que aumentaron la escala de las variaciones. El resultado es que la conformación de estrellas que vemos en esa constelación existe tan sólo en un abanico muy limitado de universos. En la mayoría de las variantes próximas de nuestro universo también hay constelaciones en el cielo, pero tienen configuraciones diferentes en cada una de ellas.
Por último, contemplemos del mismo modo el universo que nos rodea. ¿Qué captará la atención de nuestra mirada mágicamente potente? En un universo aislado, las estructuras más impresionantes son las galaxias y los cúmulos de galaxias. Pero tales objetos no tienen una estructura discernible a lo largo y lo ancho del multiverso. Donde aparece una galaxia en un universo, habrá una miríada de galaxias, con geografías muy distintas, en el multiverso. Y ello es así en todos los aspectos. Los universos cercanos sólo se parecen en ciertas características macroscópicas aproximadas, como requieren las leyes de la física, que son de aplicación a todos ellos. Así pues, la mayoría de las estrellas serán aproximadamente esféricas en cualquier parte del multiverso, y la mayor parte de las galaxias serán elípticas o espirales. Pero nada puede extenderse profundamente por universos cada vez más lejanos sin que su estructura detallada cambie de modo irreconocible. Sólo son excepciones de esta regla los escasos lugares en que el conocimiento se ha materializado físicamente. En ellos los objetos se extienden por grandes cantidades de universos sin perder por ello las características que los hacen reconocibles. Quizás la Tierra sea el único lugar así de nuestro universo, de momento. En cualquier caso, esos lugares se distinguen, en el sentido que he descrito, por ser aquellos en que se localizan los procesos —la vida y el pensamiento— que han creado las mayores estructuras distintivas que hay en el multiverso.