
FIGURA 7.1: La combinación de dos mutaciones en el gen Ultrabithorax genera una mosca con cuatro alas en vez de dos. La mosca es una pesadilla, sobre todo para los neodarwinistas.
Vimos en el primer capítulo que la teoría de la evolución —la propuesta de que todos los seres vivos de la Tierra proceden de un único ancestro— fue aceptada por la mayoría de los científicos en cuanto la publicó Darwin en 1859, mientras que el mecanismo de la selección natural nunca gozó del mismo predicamento. A finales del XIX y principios del XX, todos los científicos eran evolucionistas convencidos, pero la selección natural vivía sus momentos más bajos. Una de las razones era que muchos naturalistas creían aún que los procesos de tipo lamarckiano, basados en la herencia de los caracteres adquiridos, aportaban a la evolución un mecanismo mucho más eficaz que los ciegos y tediosos meandros de la selección natural. Nadie podía culparles. Al fin y al cabo, el propio Darwin había derivado ostensiblemente hacia el lamarckismo en sus últimos años. Pero fue el redescubrimiento de las leyes de Mendel, en 1900, el que vino a embarrar irremisiblemente la situación. Los genes eran entidades discretas, y sus mutaciones parecían tener efectos discontinuos en el organismo. Una sola mutación transformaba un guisante liso en rugoso, o en blanco el ojo rojo de una mosca, sin gradaciones ni medias tintas. Si el darwinismo era un mundo de cambio gradual y ritmos uniformes, todo en la recién nacida genética parecía discontinuo, saltarín, discreto.
De hecho, la genética fue utilizada desde el primer momento como un arma arrojadiza contra el gradualismo darwiniano. El científico británico William Bateson (1861-1926), uno de los cuatro investigadores que redescubrieron en 1900 las leyes de Mendel —y el que inventó el término genética—, ya era un saltacionista incluso antes de esa fecha, cuando publicó su influyente trabajo Materiales para el estudio de la variación (1894). Bateson concluía allí que la evolución no podía ocurrir mediante el cambio continuo de las especies, puesto que, en las plantas y animales que había observado, las estructuras aparecían y desaparecían súbitamente, o se transformaban de una en otra de forma discreta y completa. Gran aficionado a bautizar los nuevos conceptos científicos, Bateson inventó el término homeosis para referirse a esa transformación discreta y completa de un órgano en otro. Esa palabra acabó siendo su contribución más perdurable a la historia de la biología, como veremos en seguida.
Bateson fue el gran introductor de Mendel en la ciencia anglosajona. En 1900, tras reconocer honradamente que las leyes de la herencia descubiertas 34 años antes por el monje austríaco podían explicar sus observaciones mejor que sus propias teorías, se ocupó de traducir al inglés el artículo de Mendel y se convirtió en su predicador oficial en tierras británicas. Curiosamente, en la primera década del siglo XX, fue también Bateson el primero en descubrir que los genes no siempre se transmitían de forma independiente a la siguiente generación, como había formulado el monje en una de sus leyes, sino que a veces hacían ese viaje juntos. Hoy sabemos que esto ocurre cuando varios genes están en el mismo cromosoma, un hecho enormemente común que, con toda probabilidad, Mendel ya había descubierto 40 años antes pero se guardó muy mucho de publicar para no empantanar sus nítidas estadísticas. En 1905 no se sabía aún que los genes estaban en los cromosomas, y Bateson, que no supo interpretar bien sus propios datos, formuló una teoría vibratoria de la herencia que ahora no es posible recordar sin sonrojo.
Fue el estadounidense Thomas Hunt Morgan quien interpretó correctamente los datos de Bateson, para horror del inglés, que nunca aceptó que los genes pudieran estar ordenados en fila a lo largo de cada cromosoma. La caída de Bateson a manos de Morgan tiene su importancia histórica, porque marca el inicio de la hegemonía científica norteamericana que caracterizaría a todo el siglo XX recién inaugurado, y que seguimos presenciando a comienzos del XXI. A partir de 1910 y durante varias décadas, Morgan y su prodigioso equipo de la Universidad de Columbia de Nueva York utilizaron la mosca del vinagre Drosophila melanogaster y sus grandes ventajas prácticas —prolijidad, pequeño tamaño y gran velocidad de desarrollo— para fundar la genética moderna. El equipo de Morgan estableció para siempre que los genes no sólo eran las unidades de la herencia descubiertas por Mendel, sino también los motores de la evolución formulada por Darwin y los conductores que guían al óvulo fecundado —una sola célula— a través del desarrollo embrionario hasta fabricar un adulto completo.
Al igual que Bateson, Morgan interpretó inicialmente los datos de la genética, con sus saltos discontinuos y sus mutaciones bruscas, como un desafío al gradualismo darwiniano. Pero uno de sus discípulos, el ruso Theodosius Dobzhansky, fue uno de los científicos que más contribuyeron a disipar esas aparentes discrepancias. Dobzhansky se unió al grupo de Morgan en 1927 y, diez años después, tras compilar las evidencias que esos pioneros de la Universidad de Columbia iban obteniendo sobre la naturaleza de los genes, publicó Genética y el origen de las especies (1937), un trabajo de enorme influencia que convenció a varias generaciones de evolucionistas de que el efecto morfológico de las mutaciones podía ser en verdad muy pequeño, y mostró cómo la combinación de varias de ellas se bastaba para conferir a una población un repertorio continuo de variación externa enteramente compatible con el gradualismo darwiniano: un repertorio gradual de formas sobre el que podía actuar a su antojo la selección natural.
El trabajo de Dobzhansky, que se apoyaba firmemente en los modelos matemáticos del norteamericano Sewall Wright (el inventor del modelo de especiación alopátrica), obró el prodigio de compatibilizar dos ideas previamente percibidas como antagónicas, la selección natural y la genética mendeliana, una hibridación que se conoce desde entonces como teoría sintética o neodarwinismo. El postulado central de la teoría sintética, que ha sobrevivido intacto hasta nuestros días, es que, por más discretos y bruscos que sean los efectos de algunas mutaciones, la interacción entre muchas de ellas puede generar un continuo de variación en la población sobre el que actúan los graduales y parsimoniosos mecanismos de la selección natural. El aislamiento geográfico de una subpoblación hace el resto, al permitir la especiación (la generación de una nueva especie) por acumulación de cambios graduales en un sentido distinto al de población general. Para los neodarwinistas, como para los darwinistas sin más, los grandes cambios evolutivos, como la generación de nuevas especies, no necesitan mecanismos genéticos especiales: basta el cambio gradual acumulado por selección natural una generación tras otra, con la ayuda del aislamiento alopátrico de Wright desarrollado por Ernst Mayr.
Por cierto que, para dar una idea de la influencia de Dobzhansky, cabe recordar que fue su libro Genética y el origen de las especies el que logró convertir al darwinismo al mismísimo Mayr, que a principios de los años treinta sostenía todavía mecanismos lamarckianos para explicar sus observaciones sobre los pájaros de Nueva Guinea y las islas Solomon. Tras la lectura de Dobzhansky, Mayr se convirtió en el darwinista más prestigioso de la segunda mitad del siglo XX. Y hay otro punto sobre Dobzhansky que no quiero dejar escapar. Lo cito del filósofo de la evolución Michael Ruse:
Desde el principio, Dobzhansky reconoció que se dedicó a la empresa evolucionista con una misión, en su caso religiosa: la esperanza de demostrar que la evolución tiene un propósito divino y que el hombre era su producto más perfecto, la apoteosis de un proceso ascendente y progresivo. (RUSE, 2001.)
Lo que me lleva a su vez a un comentario cáustico de Lynn Margulis:
Muchas circunstancias conspiran para oscurecer los descubrimientos científicos, especialmente aquéllos que causan incomodidad sobre las normas sagradas de nuestra cultura […] Una convicción extendida aunque no formulada es la gran cadena del ser. Define la venerable posición de los humanos como el centro exacto del universo en la mitad de la cadena del ser, debajo de Dios y encima de la roca. Esta idea antropocéntrica domina el pensamiento religioso, incluso el de aquéllos que aseguran rechazar la religión y reemplazarla con una visión científica del mundo. (MARGULIS, 1998.)
Abandonemos por un momento la gran síntesis neodarwiniana, pero sin alejarnos mucho de Thomas Hunt Morgan. Ya en 1915, doce años antes de que Dobzhansky llegara desde Rusia a la Universidad de Columbia, otro de los estudiantes de Morgan, Calvin Bridges, había encontrado una mutación en Drosophila que parecía transformar parte del tercer segmento torácico (que normalmente tiene un par de pequeños órganos para estabilizar el vuelo, llamados halterios) en una copia del segundo segmento torácico (que es donde están las dos alas). El resultado es una mosca que no llega a tener cuatro alas, pero que se aproxima bastante a esa herejía taxonómica (véase la figura 7.1). La mosca es de la familia de los dípteros, un grupo que debe su nombre precisamente al hecho de tener dos alas, y no cuatro como las mariposas o las libélulas. Bridges llamó a esa mutación bithorax, y se dio cuenta de que representaba el primer ejemplo de «homeosis» causado por una mutación ocurrida ante sus ojos. ¿Recuerdan? Ése fue el término acuñado poco antes por el británico William Bateson, el gran inventor de palabras, para designar las transformaciones discretas de un órgano en otro que había observado en el mundo vegetal.
FIGURA 7.1: La combinación de dos mutaciones en el gen Ultrabithorax genera una mosca con cuatro alas en vez de dos. La mosca es una pesadilla, sobre todo para los neodarwinistas.
En 1919, cuatro años después de descubrir bithorax, Bridges encontró otra mutación «homeótica» situada muy cerca en el mismo cromosoma, a la que denominó bithoraxoid (literalmente, «cosa parecida a bithorax»). Mirando más cuidadosamente a sus dos mutantes. Bridges empezó a notar que todo el asunto revestía un carácter muy extraño. Mientras bithorax transformaba la parte anterior del halterio en una parte anterior del ala, bithoraxoid hacía lo propio con las partes posteriores de ambos órganos. Además, esta última mutación afectaba también al primer segmento abdominal (que normalmente no tiene patas), convirtiéndolo en una versión del tercer segmento torácico (que sí tiene). El resultado ahora se aproximaba a una mosca que no sólo tenía cuatro alas en vez de dos, sino también cuatro pares de palas en vez de tres, lo que ya era el colmo. Para ser un insecto hay que tener tres pares de patas. Si uno tiene cuatro pares de patas, es un arácnido.
Bridges tenía muchas otras cosas que hacer y no examinó la cuestión más a fondo, de modo que hubieron de pasar 27 años hasta que uno de sus estudiantes, Ed Lewis, retomó el problema en 1946. Así de parsimoniosa era la genética de aquellos días. Por ponerlo de una forma suave, digamos que Lewis se metió en un berenjenal cromosómico de una espesura sin precedentes y que, cuando logró zafarse a machetazos del enredo, habían pasado nada menos que 32 años más. En 1978 envió a la revista Nature el manuscrito con los resultados que había amasado después de tres décadas cruzando moscas y dibujando flechas en un papel. Los editores de Nature, que en su vida habían visto un análisis genético más farragoso, oscuro e inextricable que aquél, quisieron devolverle el manuscrito a Lewis para que intentara aclarar algunos puntos, pero el científico de Cambridge Peter Lawrence, un estrecho colaborador de Francis Crick, les disuadió con el siguiente argumento: «¡Ni se os ocurra devolverle el manuscrito! Puede tardar otros 30 años en enviarlo de nuevo». Así que el artículo apareció tal cual en 1978, titulado «A gene complex controlling segmentation in Drosophila» (Un grupo complejo de genes que controla la segmentación en Drosophila). Poca gente entendió aquel galimatías. Habían pasado 63 años desde que el mentor de Lewis, Calvin Bridges, descubrió la mutación bithorax en el laboratorio de Morgan. Me pregunto qué pensará Lewis, que todavía vive, cuando oye hablar en la tele de los «vertiginosos avances de la genética».
El problema con Lewis era que, pese a tener en sus manos durante tres décadas el más apasionante de los problemas de control genético del desarrollo animal, nunca logró interesarse a fondo por esa materia. Miembro al fin y al cabo de la escuela de Morgan, su motivación fue siempre la genética pura, y bithorax no era para él más que un modelo de estudio de la regulación genética. El propio Lewis lo reconoció en 1995 en Estocolmo, cuando recogió el premio Nobel:
Era evidente que la serie diversa de municiones del tipo bithorax albergaba la promesa notable de representar un grupo de genes, y no una serie alélica [una serie de distintas variaciones de un solo gen]. Fue por esta razón por la que fueron elegidos como objeto de estudio, y no por ninguna creencia en que nos dirían algo sobre cómo los genes controlan el desarrollo. (LEWIS, 1995.)
La plena implicación de los genes del tipo bithorax (a los que llamaremos a partir de ahora genes Hox) en el campo del control genético del desarrollo y la evolución animal debe mucho a lo que en los años setenta se llamó «la escuela de Madrid», que en realidad era un pequeño laboratorio de moscas dirigido por Antonio García-Bellido, entonces en el Centro de Investigaciones Biológicas del Consejo Superior de Investigaciones Científicas. García-Bellido y sus entonces estudiantes de doctorado Ginés Morata y Pedro Ripoll demostraron en los primeros años setenta que el cuerpo de la mosca se va dividiendo, a lo largo del desarrollo, en territorios estancos a los que llamaron compartimentos. Cada segmento, por ejemplo, está dividido desde los primeros minutos de vida del embrión en un compartimento anterior y uno posterior. Cada célula sabe en todo momento a qué compartimento pertenece, y ni ella ni sus hijas cruzan jamás la frontera entre compartimentos, pese a que ésta es invisible y no supone ninguna barrera física obvia. Estos experimentos y teorías provocaron que el codescubridor de la doble hélice del ADN, Francis Crick, se interesara en los resultados del grupo español hasta el punto de escribir una revisión sobre ellos, en la revista Science, junto con su colaborador Peter Lawrence, que había visitado el laboratorio de Madrid y conocía de primera mano el trabajo.
El laboratorio de García-Bellido pudo establecer ya entonces que cada gen Hox funciona en todas las células de cada compartimento para otorgar a éste una identidad concreta: por ejemplo, la identidad del compartimento anterior del tercer segmento torácico. De ahí que la mutación bithorax transformara la parte anterior del halterio en parte anterior del ala, mientras que la mutación bithoraxoid hacía lo propio con las partes posteriores: cada mutación afecta a la identidad de un compartimento concreto. Para un pequeño grupo de genetistas de Drosophila de varios países, resultó evidente que los genes Hox conferían la identidad a cada compartimento regulando a baterías completas de otros genes, y la biología molecular vino a darles la razón en la siguiente década.
En los primeros años ochenta, Ginés Morata se planteó la desapacible tarea de esclarecer el laberinto que Ed Lewis había depuesto en su Nature de 1978. En los alrededores de bithorax, Lewis creía haber descubierto más de una docena de genes Hox: bithorax, bithoraxoid, postbithorax, Contrabithorax, Ultrabithorax y un grupo de otros ocho genes implicados en el desarrollo abdominal, a los que llamó infraabdominal-2, infraabdominal-3 y así hasta infraabdominal-9. La relación entre estos genes era más bien tormentosa y venía regida por un enjambre de reglas extravagantes formuladas por el propio Lewis con el único propósito de cuadrar sus datos, tales como ésta: «Las mutaciones de pérdida de función de un gen dado tienden a inactivar polarmente los genes próximos más distales». O esta otra: «La pérdida de función de un gen tiende a estar acompañada de la sobreexpresión de la función asociada con el gen inmediatamente proximal».
Ginés Morata encargó los experimentos a su estudiante de doctorado Ernesto Sánchez Herrero. El trabajo implicaba la generación de decenas de nuevas mutaciones y un cuidadoso estudio de su efecto, en solitario y en múltiples combinaciones entre ellas. Pronto observaron que los resultados no encajaban con las ideas de Lewis, y que todo el enjambre genético deducido por éste podía, en realidad, reducirse a sólo tres genes bastante convencionales.
Morata dio en aquel año varias conferencias en distintos centros de investigación del mundo para anunciar que el embarullado enjambre propuesto por Lewis tras 32 años de experimentación era erróneo, y que aquella región cromosómica no tenía en realidad más que tres genes bastante normales. Lewis era ya una figura intocable, al menos en Estados Unidos, y las ideas de Morata fueron recibidas con un generalizado escepticismo. Pero ese mismo año ocurrió algo extraordinario.
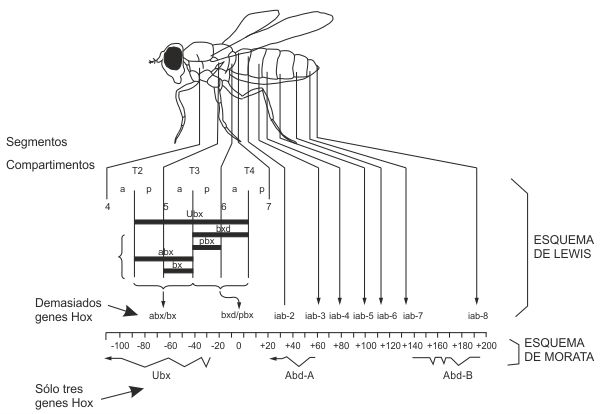
FIGURA 7.2: Ed Lewis complicó demasiado el mapa genético de los genes Hox: llamó «genes» a lo que, en realidad, eran zonas reguladoras de cada gen (arriba). Ginés Morata elucidó después el modelo correcto (abajo).
Los biólogos moleculares ya habían purificado el ADN de las regiones del genoma de Drosophila que, según habían definido Bridges, Lewis y otros genetistas, contenían los genes Hox. Estos tramos de ADN medían en conjunto cerca de un millón de bases (las letras de la doble hélice), un espacio en el que podrían caber hasta un millar de genes de tamaño pequeño. Los procedimientos eran muy tediosos en aquellos años, y rastrear esas amplias zonas de ADN hasta descartar la paja, encontrar los genes predichos por Lewis y describirlos en detalle llevaría aún mucho tiempo. Pero William McGinnis y Michael Levine, del laboratorio de Walter Gehring en Zurich, y por otro lado Matt Scotl y A. J. Weiner, en Estados Unidos, tuvieron una idea simple y eficaz que iba a acelerar enormemente las cosas.
Nadie sabía qué tipo de secuencia de ADN deberían tener los genes Hox. Pero, puesto que todos esos genes parecían hacer cosas similares —definir la identidad de uno u otro compartimento a lo largo del cuerpo de la mosca—, era bastante probable que los genes tuvieran cierto parecido entre sí. Detectar zonas de secuencia parecida en el ADN es muy sencillo: como consecuencia de la complementariedad de bases, tienden a pegarse entre sí en ciertas condiciones. Así que McGinnis, Levine y los demás decidieron dar un palo de ciego. Sólo les llevaría un par de días, y no tenían nada que perder. Simplemente, partieron en centenares de pedazos todo el ADN que tenían purificado, y buscaron si algún pedazo se pegaba a algún otro, sin ninguna idea preconcebida. ¡Bingo! Tal y como habían esperado, había una pequeña secuencia de ADN, de unas 180 hases de longitud, que se pegaba en varios sitios. La llamaron homeobox, que en la jerga molecular viene a querer decir «secuencia de ADN que suele aparecer en los genes homeóticos». ¿Ven lo que dije antes? Fíjense hasta dónde llegó la palabra inventada por Bateson, homeosis, su más perdurable aportación a la ciencia. El término Hox, por cierto, no es más que una contracción de la palabra homeobox.
En la región de los alrededores de bithorax, donde Lewis había creído ver una docena de genes, McGinnis y Levine sólo encontraron tres homeoboxes. Ese solo hecho, aun antes de saber qué era exactamente la homeobox, convenció a todo el mundo (excepto a Lewis) de que Morata tenía razón. El modelo de «los tres genes bastante normales» se publicó en 1985 en Nature con todos los honores, y se ha convertido desde entonces en uno de los trabajos españoles más citados por las revistas científicas internacionales. (Drosophila tiene otros seis genes Hox, pero están en un bloque separado, alejado de bithorax.)

FIGURA 7.3: Ginés Morata (de pie) asegura haber pescado ese desconcertante salmón de 6,2 kilos.
El descubrimiento de la homeobox fue tan estimutante para todo el mundo que McGinnis y Levine pudieron publicarlo en Nature inmediatamente, aquel mismo año de 1984, sin siquiera haber determinado qué significaba esa secuencia de ADN, algo que sería por completo inimaginable en nuestros exigentes días. Pero pocos días después de aparecer ese artículo, el biólogo molecular Carlos Cabrera llegó a Madrid desde el laboratorio de David Hogness, en California, con cotilleos mucho más frescos. Harto de que le pararan por los pasillos para pedirle noticias. Cabrera decidió dar un seminario informal para contar que la homeobox representaba un módulo presente en todas las proteínas Hox (las proteínas fabricadas por los genes Hox) que permitía a éstas unirse al ADN, lo que indicaba que la función de los genes Hox era regular a otros genes.
—¡Toma, pues vaya novedad! —bramó desde su asiento García-Bellido.
—Bueno —se disculpó Cabrera—, todavía hay por ahí quien sostiene que son proteínas de membrana.
—¡Memeces! —dijo García-Bellido, y ahí se acabó el seminario.
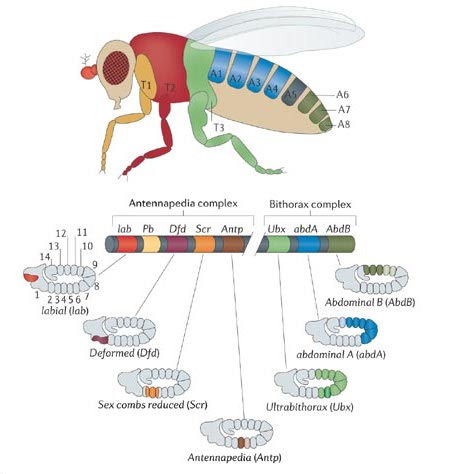
FIGURA 7.4: Lewis descubrió que los genes de la fila Hox están dispuestos en el cromosoma en el mismo orden que las partes del cuerpo que define cada uno de ellos. El embrión de Drosophila está doblado sobre si mismo: por eso los últimos segmentos del cuerpo se sitúan encima de los primeros.
Decenas de laboratorios de todo el mundo empezaron a trabajar con la homeobox, y algunas de las mayores sorpresas de la biología del siglo XX se encadenaron a velocidad de vértigo. La homeobox no es una peculiaridad de Drosophila, sino que aparece sistemáticamente en todos los animales, y ello permitió purificar los genes Hox de cualquier especie directamente, sin necesidad de análisis genético previo (no fue necesario un Ed Lewis trabajando 32 años por cada nueva especie). Cada animal tiene aproximadamente una decena de genes Hox, y siempre aparecen dispuestos en fila. La única excepción, paradójicamente, es Drosophila, donde la fila primordial aparece dividida en dos bloques. Pero, hecha esta salvedad, el orden de los diez genes es siempre el mismo en todas las especies. Ed Lewis había demostrado en Drosophila otro hecho increíble: que el orden de los genes Hox en el cromosoma es el mismo que el orden de las partes del cuerpo que cada gen define. A la izquierda los genes que especifican la cabeza, en el centro los del tronco y a la derecha los del abdomen. Esto se llama colinearidad.
Pues bien, cuando los genes se pudieron inactivar artificialmente en ratones y otras especies, se comprobó que guardaban el mismo principio de colinearidad que en la mosca. Para colmo, los genes son intercambiables entre especies. El gen Hox llamado Deformed (uno de los primeros de la fila, véase figura 7.5) especifica en la mosca una parte de la cabeza, y su equivalente en otras especies también especifica parte de la cabeza de un sapo, un ratón o un ser humano. El último gen Hox de la fila se llama caudal, y define la zona anal en todas las especies. Un gen Hox humano es capaz de curar a una mosca que tenga mutado el gen equivalente. Ah, y lo que el gen humano le restaura a la mosca mutante es una cabeza de mosca, no una cabeza humana: los genes Hox no hacen estructuras, sólo seleccionan una u otra estructura entre las disponibles para cada especie. La fila de genes Hox se ocupa de que las estructuras propias de una especie estén en el lugar correcto a lo largo de un eje.
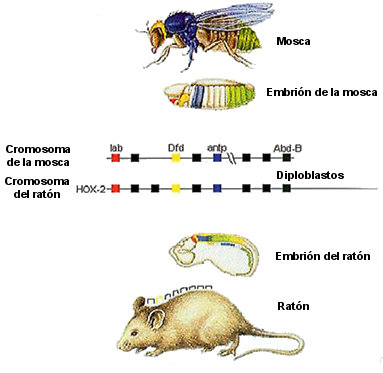
FIGURA 7.5: La mayor sorpresa de la biología de la segunda mitad del siglo XX: la fila Hox existe en todos los animales (aquí, la mosca y el ratón), y se dedica a lo mismo en todos ellos.
Todo lo que he expuesto en el último párrafo se ha ido desplegando en los últimos 15 o 20 años ante los ojos exorbitados de los genetistas. Se trata, en mi opinión, del conjunto de hechos más sorprendente y enigmático que la genética ha descubierto en toda su historia, porque revela que toda la deslumbrante diversidad animal de este planeta, desde los ácaros de la moqueta hasta los ministros de cultura pasando por los berberechos y los gusanos que les parasitan, no son más que ajustes menores de un meticuloso plan de diseño que la evolución inventó una sola vez, hace unos 600 millones de años. Y que, sin embargo, es tan eficaz y versátil que sirve para construir casi cualquier cosa que uno quiera imaginar, nade, corra, vuele o resuelva ecuaciones diferenciales. Nadie, absolutamente nadie, se hubiera imaginado una cosa semejante hace 20 años, no digamos ya en tiempos de Darwin.
La explosión cámbrica sigue siendo un gran problema, pero ya no es el gran problema. La aparición de todos los bauplanes actuales durante aquel corto periodo de 10 millones de años puede seguir pareciendo un fenómeno desconcertante, pero la verdad es que palidece frente al enigma al que nos enfrentamos ahora. Si el precursor de todos los animales de la explosión, un ser precámbrico desconocido al que hemos denominado Urbilateria, estaba ya dotado de un meticuloso mecanismo de diseño basado en una fila de genes Hox, es decir, en un sistema genético especializado en organizar cualquier tipo de cuerpo del futuro, un sistema tan complejo, eficaz y versátil que ha permanecido universalmente incuestionado durante los 600 millones de años subsiguientes, ¿de dónde demonios salió Urbilateria?