
¿Cuánta energía necesita un ciclista para mover su bicicleta? ¿Cómo genera su cuerpo la energía necesaria? La física newtoniana aporta la respuesta a la primera pregunta. Contestar a la segunda, requiere una incursión, siquiera muy somera, por los dominios de la bioquímica. Pero además, una reflexión sobre la energía reclama la ayuda de las dos leyes fundamentales de la termodinámica. Porque están en juego los conceptos de trabajo, potencia y energía y asoma entre líneas el sutil concepto de entropía, tan frecuentemente citado y tan escasamente comprendido.
En lo que sigue, he pretendido, simplemente, recuperar la memoria sobre algunas cuestiones de cultura general, relacionadas con la química de la vida, que muchos hemos sabido alguna vez y que ninguno deberíamos haber olvidado del todo.
Ya se ha recordado en el apartado 4, que el ciclista para desplazarse con su montura realiza un trabajo que, genéricamente, se define como la fuerza que actúa sobre un cuerpo por la distancia que éste recorre en la dirección de la fuerza. Las unidades que habitualmente se utilizan para cuantificar el trabajo o la energía son el julio, el kilowatio-hora y la kilocaloría.
La caloría es la energía necesaria para elevar 1 grado centígrado la temperatura de 1 gramo de agua. El inglés James Prescott Joules (1818-1889), que trabajó como director de una fábrica de cerveza, y fue científico en sus ratos libres, demostró con un sencillo e ingenioso experimento que la caloría equivale a 4,186 julios, denominación que en castellano proviene del apellido distorsionado de nuestro admirado científico aficionado. La relación entre las tres unidades más habituales de trabajo y energía, son
1 kilowatio-hora = 1.000 julios/s · 3.600 s = 3,6 · 106 julios
1 kilocaloría = 4.186 julios = 4.186 julios · 1 kWh / 3,6 · 106 julios = 1,16 · 10–3 kWh
La potencia, por su parte, es el trabajo o energía por unidad de tiempo y suele medirse en watios (julios/segundo) o en alguno de sus múltiplos, como el kilovatio (103 W) o el megavatio (106 W). Aunque también es habitual, en el mundo del motor, utilizar el caballo de vapor, CV o HP (horse-power en la terminología anglosajona), siendo la equivalencia
1 CV ≈ 736 watios
La denominación de watio proviene del apellido del ingeniero inglés, James Watt (1736-1819), que en la segunda mitad del siglo XVIII acabó de poner a punto la máquina de vapor, que al transformar la energía calorífica en energía mecánica, hizo posible la sustitución de la tracción animal por la mucho más eficiente tracción mecánica. La invención de la máquina de vapor fue capital para impulsar la revolución industrial inglesa, que fue contemporánea de la revolución francesa, y que, de otra manera, más para bien que para mal, aunque de todo haya habido y haya en la viña del Señor, contribuyó a transformar el mundo.
El cuerpo humano es una máquina, fascinante y compleja, que transforma la energía química, que proviene de los alimentos con los que se nutre, en la energía mecánica que, en parte, utiliza el cuerpo humano para mover sus músculos y, en mucha mayor medida, se hace calor, en coherencia con el 1er principio de la termodinámica.
Los complejísimos procesos metabólicos que en los seres humanos hacen posible el movimiento, y por tanto, el milagro de la vida, están muy estudiados y se conocen con gran precisión. Aquí recordaremos solamente, somera e imprecisamente, algunos aspectos de dichos procesos.
El «1er Principio de la Termodinámica» nos dice que la variación de energía interna de un sistema, el del cuerpo del ciclista y su entorno ambiental, es igual al trabajo externo que realiza más el calor que genera
∂E = W + Q
∂E = variación de la energía interna
W = trabajo externo realizado
Q = calor generado
Lo que nos viene a recordar, también, que «la energía ni se crea ni se destruye, solo se transforma».
Desde una perspectiva energética al cuerpo humano se le puede considerar muy poco eficiente. Sólo un 24% aproximadamente, de la energía que le aportan los alimentos se pueden convertir en trabajo mecánico. Una buena parte del resto se dedica al mantenimiento de las funciones fisiológicas, asociadas al metabolismo basal: respiración, circulación sanguínea, digestión, excreciones, mantenimiento de la temperatura corporal, crecimiento, reconstrucción, etc.
La energía solo se transforma, pero algunas formas de energía pueden ser más aprovechables que otras. De la posibilidad o imposibilidad de hacer un uso útil de la energía trata el 2º Principio de la termodinámica, que nació años después del primero, para aportar las respuestas que éste no podía dar. Tuvo diversos enunciados. Como los que formularon Kelvin, Clausius o Carnot, orientados al análisis del rendimiento de los motores de vapor y térmicos.
Con carácter más general, el enunciado del 2º Principio nos dice que durante un proceso irreversible la entropía total de un sistema y su entorno aumenta siempre. La entropía es una función termodinámica que mide el desorden de un sistema. La entropía S —como la presión, V, la temperatura, T, y la energía interna, Eint— es una función de estado de un sistema: el cambio de un estado inicial a otro final no depende del proceso, tan solo cuenta cuál es el origen y cuál es el estado final. Por ello, en la entropía, como en la energía potencial y cinética, lo que es importante, es lo que cambia y no los valores absolutos. Se puede definir, dicho cambio diferencial, con referencia al calor, Qrev, absorbido en un proceso reversible, mediante la expresión
∂S = ∂Qrev / T
En un proceso irreversible la energía también se conserva, pero parte de ella no está disponible para hacer trabajo útil.
Consideremos un bloque de masa, m, que colocado a una altura h del suelo acumula una energía potencial m · g · h que podría transformarse en trabajo útil. Pero si el bloque cae y se produce un choque totalmente inelástico contra el suelo, se habrá perdido dicha posibilidad. Con el impacto habrá aumentado la energía interna de las moléculas que conforman el bloque de masa m. La energía perdida, m · g · h, sería igual a T · ∂S, siendo ∂S el aumento de la entropía del sistema y T la temperatura absoluta en grados Kelvin (recordamos que –273° C es la temperatura del cero absoluto, y por tanto una temperatura de 20° C equivale a 293° K). Si la masa fuese de 10 kg, la altura de la caída de 1 m y, como consecuencia del choque, la temperatura inicial del sistema (20° C) no se modificase prácticamente la entropía habría aumentado en el valor
∂S = m · g · h / T = 10 kg · 9,8 m/s2 · 1 m / 293° K = 98 julios / 293° K = 0,33 julios/° K
El ciclismo es obviamente un proceso irreversible. Como ya hemos comentado, solamente una parte de la energía que genera el cuerpo del ciclista, del orden del 24%, sirve para mover la bicicleta. Aunque no se debería considerar como energía inútil la que hace posible el funcionamiento de su cuerpo. En cualquier caso, es evidente que parte de la energía se convierte en el sudor que desprende el ciclista, y que se pierde también energía en los rozamientos de los componentes de la bicicleta que se mueven y en los que se producen entre las ruedas y el pavimento. Los ruidos y los susurros que acompañan al ciclista en su pedaleo es la música de la políglota entropía, su contribución al incremento del desorden del universo.
La energía necesaria para el movimiento de un ser humano, en general, y del cuerpo del ciclista, en particular, proviene de los nutrientes de los alimentos que consumimos:
Como los diferentes tipos de nutrientes tienen misiones específicas o complementarias, una dieta equilibrada debería aportar suficientes cantidades de unos y otros. Algunas recomendaciones dietéticas apuntan a porcentajes del 50% o 60% de hidratos de carbono, del orden del 30 al 35% de grasas y de 10 al 15% de proteínas.
Los hidratos de carbono [C6 (H2O)6]n, con n = 1 para los monosacáridos como la glucosa o la fructosa, n = 2 para los disacáridos como la sacarosa o la lactosa y con valores de n mucho más elevados en los polisacáridos, como el glucógeno o el almidón, se encuentran en los cereales, legumbres, leche, frutas y verduras y en los dulces que consumimos.
Los lípidos —triglicéridos y ácidos grasos— son compuestos orgánicos insolubles en agua, constituidos por C, H, O. Los proporcionan la mantequilla, el aceite, la carne, el pescado graso, los huevos y los productos lácteos.
Las proteínas están constituidas por aminoácidos muy diversos, con presencia del nitrógeno en proporciones del 16%. Son suministrados, en proporciones muy variables, por la carne, el pescado, los quesos, la leche, los huevos, los cereales y las legumbres y por las frutas y verduras.
A modo de referencia, se pueden recordar los aportes de kilocalorías que proporcionan 100 gramos de algunos alimentos (ver tabla adjunta).
|
Alimento |
kcal (100 g) |
|
Alubias |
300 |
|
Carne magra |
100-150 |
|
Chocolate |
500-600 |
|
Hígado |
100-150 |
|
Huevos |
100-150 |
|
Leche |
50-100 |
|
Mayonesa |
750 |
|
Merluza |
50-100 |
|
Naranja |
30-40 |
|
Queso |
300-400 |
|
Pan |
200 |
|
Patata |
100 |
|
Tocino |
700 |
La energía que nos aportan los alimentos que consumimos no suelen necesitarse hasta algún tiempo después de haberlos ingerido. Por ello, el cuerpo humano, la almacena en forma de energía potencial química. Como ocurre, en otro contexto, con las baterías que alimentan los motores eléctricos. O como sucede, de otra manera, con la energía acumulada en el agua de los embalses asociados a centrales hidroeléctricas.
En los procesos aeróbicos —los más habituales en los glúcidos y los únicos que pueden utilizar los lípidos y las proteínas— las transformaciones metabólicas que movilizan los músculos, se realizan con la colaboración imprescindible del oxígeno contenido en el aire que respiramos. La energía proveniente de los glúcidos se puede activar, también, mediante procesos anaeróbicos que no utilizan el oxígeno proveniente de la respiración porque para su transporte y asimilación por las fibras musculares se requiere un tiempo del que no se dispone. Ni las urgencias que demandan algunos movimientos musculares, casi instantáneos, ni las elevadas potencias que puntualmente requieren algunos deportes de alta competición, son compatibles con los sosegados procesos aeróbicos, que son los más habituales tanto en el deporte como en la vida normal, y a los que nos referiremos, en primer lugar.
El oxígeno que porta la sangre y proviene del aire respirado interviene en la pausada transformación de la energía química almacenada en el cuerpo, en calor y en el trabajo que hace contraer los músculos. A modo de ejemplo, la combinación de 100 gramos de glucosa (C6 H12 O6) con 75 litros de oxígeno (O2) produce otros tantos litros del famoso CO2 —de los que la respiración animal es un productor destacado— 60 litros de agua (H2O), y libera 384 kilocalorías de energía.
C6H12O6 (100 gr) + 6 O2 (75 litros) = 6 CO2 (75 litros) + 6 H2O (60 litros) + 384 kilocalorías
Un indicador muy valioso de la energía que se genera en estos procesos es el denominado «equivalente calorífico del oxígeno». En el caso analizado de la glucosa, las kilocalorías producidas por litro de oxígeno inhalado serían:
384 kilocalorías / 75 litros de oxígeno = 5,1 kilocalorías/litro de O2
Este índice es análogo al que se produce en la transformación de grasas y proteínas, según se muestra en el cuadro siguiente en el que figura, además, la energía asociada a cada gramo de estas substancias. Obsérvese que las grasas, que no contienen agua y están, por ello, muy concentradas, son, energéticamente, muy eficientes: generan 9,3 kilocalorías por gramo frente a las poco más de 4 de las proteínas y de los hidratos de carbono.
|
Alimento |
Equivalente calorífico de oxígeno (kilocalorías/litro de O2) |
Energía media metabolizable por gramo (kcal/gramo) |
|---|---|---|
|
Hidratos de carbono |
5,1 |
4,1 |
|
Grasas |
4,8 |
9,3 |
|
Proteínas |
4,5 |
4,2 |
En realidad, las proteínas generan mayores niveles de energía pero, al contrario de lo que sucede con las grasas y los hidratos de carbono, pierden una parte, del orden de 1,25 kcal/g que están asociados con el amoníaco y la urea que se elimina por la orina.
Son incontables las investigaciones que se han llevado a cabo para estimar el volumen de oxígeno, VO2, que consumen los seres humanos para generar la energía que necesitan según el tipo de actividad al que se dediquen. En el cuadro adjunto se incluyen algunos datos de referencia. En la última columna se ha indicado la potencia útil disponible, también en watios, supuesto que el rendimiento de su organismo sea del 24%.
|
Tipo de actividad |
Consumo de oxígeno l/m · (∂VO2/∂t) [mililitros / minuto · kg] |
Potencia metabólica para una persona de 65 kg (kcal/h) |
Potencia metabólica para una persona de 65 kg (watios) |
Potencia útil (P = 24%) (watios) |
|---|---|---|---|---|
|
Reposo |
3,5 |
70 |
80 |
19 |
|
ligera[1] |
10 |
200 |
230 |
55 |
|
Moderada |
20 |
400 |
460 |
110 |
|
Intensa |
30 |
600 |
700 |
144 |
|
Extrema |
70 |
1.400 |
1.627 |
390 |
El oxígeno es indispensable para movilizar los músculos que permiten a los pulmones inhalar el aire que respiramos y al corazón bombear la sangre que lo transporta. El oxígeno es el soporte indispensable de la vida. Un ser humano puede vivir unos días sin beber y algunas semanas sin comer, pero tan sólo unos escasos minutos sin respirar.
En coherencia con los valores del cuadro, el consumo de oxígeno de un ciclista profesional, en una exigente competición y durante un tiempo limitado, podría alcanzar los 70 ml/min·kg, lo que le permitiría generar una potencia corporal de 1.400 kcal/hora (1.627 W) y ejercer sobre la bicicleta una potencia útil de 0,24 · 1.627 ≈ 390 W. Como contraste una persona mientras duerme —siempre que no se trate del republicano «sueño eterno»— necesitaría 20 veces menos de oxígeno, su potencia metabólica se reduciría en idéntica proporción hasta 70 kcal/h (80 W), limitándose su potencia útil a 19 W aproximadamente.
Un varón de complexión normal que duerma 8 horas y dedique las 16 restantes a actividades sedentarias, necesitaría diariamente entre 2.000 y 3.000 kcal de las que unas 1.700 corresponden a las exigencias del metabolismo basal.
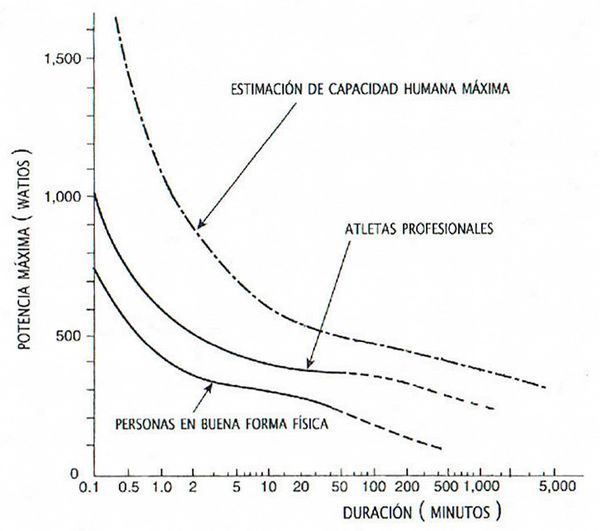
Por otra parte, la máxima potencia energética que puede generar el cuerpo de un atleta depende del tiempo que dure el esfuerzo que ha de realizar. Los especialistas de la NASA, que han investigado a fondo el comportamiento del cuerpo humano, han establecido unos rangos de valores en función de la duración del ejercicio para personas en buena forma física y para atletas profesionales bien preparados. En el gráfico adjunto, el tiempo en abscisas se representa a escala logarítmica.
Cuando batieron el récord de la hora, los registros de potencia de pedaleo de Eddy Merckx, de nuestro Miguel Induráin, o de Chris Boardman, indicaban valores de 450 a 500 W, a lo que corresponderían fuerzas aerodinámicas y de rozamiento del orden de los 30 N para velocidades de desplazamiento próximas a los 15 m/s (30 N · 15 m/s = 450 W). En otros deportes más explosivos, la potencia necesaria durante los escasos segundos o décimas de segundo que puede durar el esfuerzo, puede alcanzar los 4.000 watios, que serían de origen anaeróbico.
Se puede representar también la relación existente entre la velocidad de la bicicleta y la potencia metabólica que necesita generar el cuerpo del ciclista para desplazarse, en posición inclinada sobre la bicicleta o en una posición erguida, menos eficiente. Lógicamente, el VO2,máx será específico para cada persona, y dependerá de su capacidad pulmonar, de la eficiencia de su sistema cardiovascular para captar oxígeno y transportarlo por el fluido sanguíneo, así como por la eficacia de los tejidos musculares para almacenar y utilizar el oxígeno aportado por la sangre.
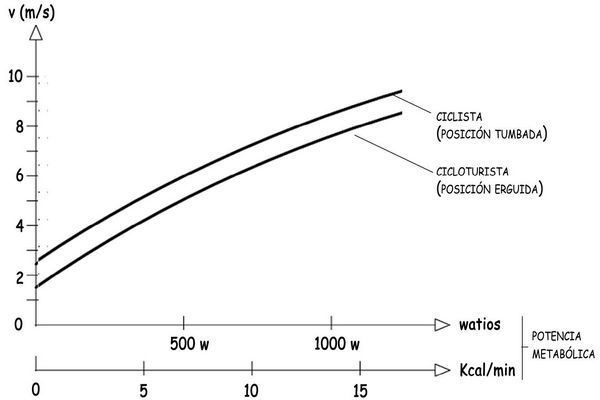
Una fórmula para estimar, en una primera aproximación, el valor de VO2,máx, en mililitros/min·kg, para un varón de t años de edad y constitución normal podría ser:
VO2,máx = 50 – 0,4 · (t – 25)
En atletas bien entrenados estos valores pueden ser muy superiores, dependiendo, por otra parte, del tipo de deporte practicado. Los índices VO2,máx de muchos ciclistas pueden superar los 70. Miguel Induráin en sus mejores tiempos parece que alcanzaba los 88 y Greg Lemond, menos corpulento, superaba los 92.
No es éste lugar inadecuado para recordar, por otra parte, que la bicicleta, con el ciclista como motor, está considerada como uno de los sistemas con mayor eficiencia energética. Estimemos, en efecto, la energía que necesitaría un ciclista de 65 kilogramos de masa para recorrer en unos 70 s la distancia de 1 km a la velocidad de 14 m/s. Las fuerzas aerodinámicas y de rozamiento, opuestas al movimiento podrían valer 30 N aproximadamente, y la potencia requerida será, por consiguiente W = F · v = 30 N · 14 m/s = 420 W. La energía que habría necesitado por gramo de masa sería
420 julios/s · 70 s/km / 65.000 gr = 0,45 julios/km · gr
Como contraste, a un caminante le podría corresponder un índice entre tres y cuatro veces mayor
100 watios · 1.000 s/km / 65.000 gr = 1,54 julios/km · gr
Cifras también superiores parece que resultan para una vaca, un caballo, o incluso para un automóvil. En alguna publicación se atribuye a los salmones índices de 4, a los conejos de 40, a las abejas de 130 y a los ratones, menos mal, superiores a 400.
Al recordar, por otra parte, estos valores que se suelen citar para entronizar la bicicleta como medio de transporte, tampoco se debe olvidar que la tracción animal, como productora de energía, es muy poco eficiente.
Hace pocos años, en la oscurecida Barcelona, se instalaron algunos artificiosos árboles navideños, revestidos de bombillas, que se encendían con la energía que generaban los ciudadanos que se prestaban a mover los pedales de unas bicicletas estáticas. La ocurrencia no produjo, claro está, los efectos que pretendieron sus desnortados promotores y en años posteriores no se volvió a repetir. Pero no está de más recordarlo aquí, porque la potencia suministrada por los ingenuos y esforzados ciudadanos que se prestaron a dar a los pedales, pudo ser de unos 100 watios. Si suponemos que se trataba de un mileurista, al que podríamos asignar un coste humano de 10 €/hora (imaginemos lo que hubiese supuesto la participación de un controlador aéreo…), el precio del kilowatio-hora generado sería de
10 euros/h / 0,100 kW = 100 euros/kWh

Este coste es mil veces superior al que supone la producción de la energía en centrales hidroeléctricas —las más eficientes—, térmicas, nucleares, eólicas y es muy superior, incluso, a la generada en parques fotovoltaicos o solares. Aquella ocurrencia ecologista era, sin saberlo y sin desearlo desde luego sus promotores, un merecido homenaje a la ingeniería.
En el mismo sentido conviene observar, también, que el extraordinario esfuerzo que realiza un ciclista profesional en alguna etapa contrarreloj de las que deciden un Tour de Francia, y duran una hora, supone un consumo de energía del orden de 400 W · 1 h = 0,40 kWh. La que se necesita para tener encendidas 4 bombillas de 100 watios durante 1 hora, con una repercusión en la factura de la luz inferior a 0,04 €.
Análogamente, la ascensión, en un muy meritorio tiempo de media hora 1.800 s, de una pendiente que salvase un desnivel de 1.000 metros, por un ciclista que con su bicicleta tuviese 70 kg de masa, supondría (sin contar las pérdidas por rozamiento) un consumo energético del orden de 70 kg · 10 m/s2 · 1.000 m = 700.000 julios, esto es, una potencia de
700.000 julios / 1.800 s = 388 watios.
La energía consumida en la ascensión sería, por tanto,
0,388 kW · 0,5 horas ≈ 0,20 kWh
y el coste de la energía eléctrica correspondiente sería inferior a 2 céntimos de euro.
En los próximos párrafos, con el corazón en la mano, recordaremos —es cultura general— cuál es su tan bien conocida y poco romántica misión principal y cuáles son algunas de sus características. Y nos referiremos también a la sangre que es su razón de ser.
El cuerpo de un varón contiene unos 75 mililitros de sangre por kilogramo, entre 5 y 6 litros en total. Un 15% menos el de las mujeres. Su corazón, que tiene unos 260 gramos de masa y el tamaño de un puño cerrado, hace circular la sangre por su organismo. Late unas 72 veces por minuto cuando está sosegado, y en cada latido impulsa unos 100 mililitros, de manera que, en menos de sesenta segundos, la totalidad de la sangre recorre por completo el cuerpo humano.
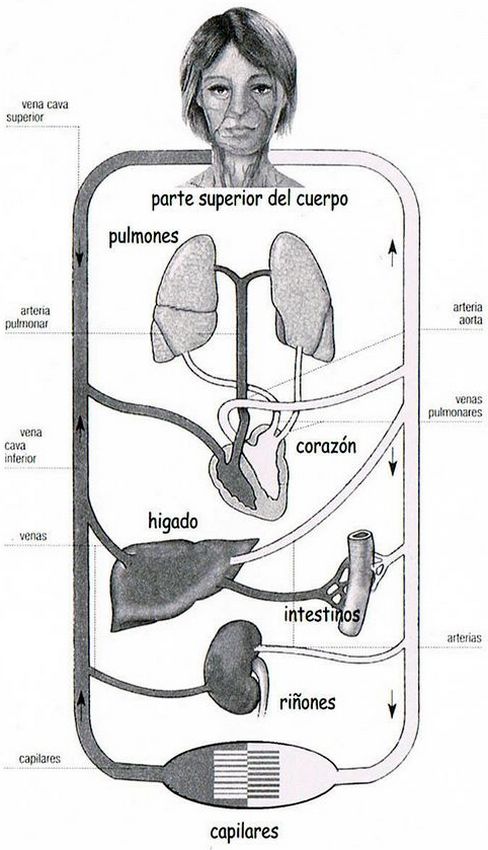
El corazón distribuye la sangre al organismo, por dos circuitos separados, y está dotado, para ello, de dos aurículas y, separadas por sendas válvulas unidireccionales, de dos ventrículos. La aurícula izquierda recibe la sangre oxigenada proveniente de los pulmones. De allí pasa al ventrículo del mismo lado que al contraerse en el movimiento de sístole, la impulsa a través de la aorta a las arterias que la difunden por todo el organismo. La aurícula derecha, a través de las venas cavas superior e inferior, recibe la sangre pobre en oxígeno, y en su diástole, la impulsa hacia los pulmones, donde se nutre de oxígeno antes de volver al corazón para iniciar un nuevo recorrido por las arterias, y capilares que la distribuyen por los tejidos corporales.
Cada pulsación, cuando el ritmo es de 72 por minuto, requiere un tiempo de unos 0,8 segundos, la mitad aproximadamente para la sístole (0,1 segundos para la recepción de la sangre y 0,3 para su expulsión) y los 0,4 segundos siguientes para la diástole. La sístole provoca una presión en las paredes arteriales del orden de 120 mm de mercurio (Hg) que en la diástole se reduce a 80 mm. Casi todo el mundo sabe que cuando nos medimos la tensión, el rango de normalidad, en general, se sitúa entre valores de 12 y 8, con variaciones por arriba, en individuos hipertensos, y por abajo, en los hipotensos, que pueden ser manifestación de posibles patologías. Menos gente recuerda que realmente dichos valores de la presión arterial están expresados en decenas de milímetros de mercurio. El científico italiano Evangelista Torricelli, que fue discípulo de Galileo y murió en Florencia en 1647, en sus experimentos pioneros utilizó un metal líquido, el mercurio, que es 13,6 veces más denso que el agua y mucho más sensible a las variaciones de temperatura que el vidrio. Por eso, aún hoy los termómetros con los que estamos más familiarizados constan de un tubo graduado de cristal que encierra una columna de mercurio, con su característica tonalidad plateada.
La presión sanguínea de 12, corresponde a la producida por una columna de mercurio de 120 milímetros de altura y es equivalente a la que ejercería una de agua (o de sangre que tiene análoga densidad) de 13,6 · 120 = 1.630 mm.
La presión arterial no es la misma en cualquier lugar del cuerpo. Así, en los pies de una persona erguida, a la presión generada por la sístole del corazón, situado, por ejemplo, a 1,20 m de altura, habría que añadir la creada por el propio peso de la sangre en las arterias, y podría alcanzar la correspondiente a 1.630 + 1.200 = 2.830 mm de altura equivalente de agua. Por esta razón se suele medir la tensión arterial en la zona del antebrazo próxima al codo que se encuentra a la altura del corazón y en situación de reposo. Es éste también el motivo por el que se recomienda que el cuerpo no pase bruscamente desde una posición tumbada a otra erguida, porque la sangre para acceder al cerebro necesitará un mínimo de tiempo.
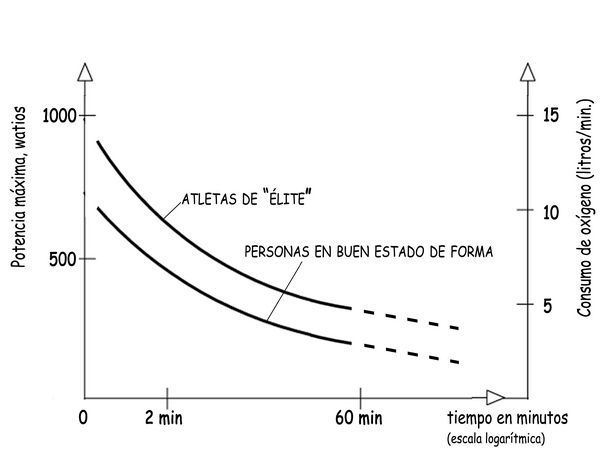
El ritmo de 72 pulsaciones por minuto en un cuerpo sometido a actividades ligeras corresponde, como hemos visto en un cuadro anterior, a un consumo de oxígeno de unos 10 ml/min·kg, y a una potencia metabólica de unos 230 W. Cuando se intensifica el esfuerzo, dichos valores se incrementan más o menos linealmente y alcanzan, por ejemplo, consumos de O2 de 30 ml/min·kg para potencias de unos 700 W que pasan de los 1.600 cuando el consumo de O2 alcanza 70 ml/min·kg.
Para alcanzar estos últimos niveles de esfuerzo las pulsaciones del corazón se multiplican también casi por tres y pueden alcanzar valores máximos que, en primera aproximación, se pueden estimar en 220 menos la edad de la persona en años: a un joven cincuentón, le corresponderían unas 170 pulsaciones/minuto. Buen lugar éste para recordar, también, que «el corazón que vive poco, late deprisa». Lo que ocurre, por ejemplo, con las aves. Este incremento de pulsaciones pretende, lógicamente, que el corazón bombee más sangre al sistema arterial y el organismo disponga de más oxígeno para transformar aeróbicamente la energía química almacenada en energía mecánica. Para lograr este imprescindible objetivo, el ritmo de la respiración también se acelera, triplicándose, como los latidos del corazón, en relación con el habitual de 15 respiraciones por minuto.
La sangre es el fluido vital. Transporta el oxígeno y los nutrientes que necesita el cuerpo humano, así como las sustancias necesarias para los metabolismos celulares. El 55% de la sangre está constituida por plasma, que tiene un 90% de agua, un 8% de proteínas y electrolitos diversos. Envueltos por el plasma se encuentran tres corpúsculos en suspensión: los glóbulos rojos (denominados también hematíes o eritrocitos) que son los encargados de transportar el oxígeno, los glóbulos blancos, o leucocitos, componentes esenciales del sistema inmunitario y las plaquetas que colaboran en los procesos de coagulación sanguínea. En cada mm3 de la sangre de un varón existen entre 4,4 y 5,4 millones de glóbulos rojos (un 10% menos en las mujeres), de 4.000 a 10.000 glóbulos blancos y 200.000 a 400.000 plaquetas.
Los glóbulos rojos se forman fundamentalmente en las médulas óseas. Tardan 4 o 5 días en hacerlo y viven unos 120 días. Tienen geometría de lentilla bicóncava entre 6 y 9 micras (10–6 m) de diámetro, para favorecer los intercambios gaseosos a través de su superficie, y disponer de la deformabilidad que requiere su tortuoso viaje por conductos circulatorios que a veces no sobrepasa las pocas micras de diámetro. La hemoglobina es una proteína que se encuentra en el interior de los glóbulos rojos, en una proporción de unos 15 gramos por cada 100 mililitros de sangre. Se combina con el oxígeno formando la oxihemoglobina que fluye por la sangre para, tras llegar a su destino, disociarse de nuevo liberando el oxígeno transportado. La hemoglobina tiene la geometría aproximada de una esfera de 5,5 nanómetros (5,5 · 109 m) de diámetro. A la salida de los pulmones, se encuentra saturada al 96% de O2, mientras que cuando retorna sólo lo está al 64%: sólo un tercio del oxígeno que transporta la sangre se queda en los tejidos corporales, unos 6,5 ml de O2 por cada 100 ml de sangre. La hemoglobina, cuando retorna con la sangre, transporta también el hidrógeno, H–, y el CO2, generados en los procesos metabólicos, hasta los pulmones y los riñones que los eliminan por la respiración y por la orina.
La esencial importancia de la sangre y del oxígeno en el comportamiento de un cuerpo humano justifica que los entrenamientos de los deportistas estén orientados a incrementar la capacidad impulsiva de su corazón, aumentar los porcentajes de oxígeno que pueda incorporar al sistema sanguíneo, mejorar la eficiencia en su captación y optimizar los procesos de transformación metabólicos para poder disponer de mayor energía y poder utilizarla más rápidamente. Con tal finalidad se suelen programar ejercicios que provoquen diferentes niveles de pulsaciones, durante periodos determinados seguidos de otros de descanso. En la preparación de atletas para la alta competición, la programación de tandas de entrenamiento y el seguimiento de resultados, es lógicamente fundamental. Y la colaboración de los profesionales de la preparación física esencial. Contribuirán a mejorar rendimientos, minimizar el riesgo de lesiones, y reducir los tiempos de recuperación si llegan a producirse. El deporte, por otra parte, realizado en forma moderada y controlada, puede ser una actividad fundamental para preservar la salud y mejorar el bienestar de quienes los practican. Aunque el cuerpo humano sea un gran desconocido, incluso para quienes habitamos en él, los progresos científicos han hecho posible que aumente notablemente nuestra esperanza de vida. Vivimos más años y podemos vivirlos saludablemente para alcanzar un objetivo que podría ser deseable para todos: morir a edad muy avanzada, llenos de salud. Conocer nuestro cuerpo y ejercitarlo sensatamente nos ayudará a lograrlo.
Hasta aquí este relato se ha referido exclusivamente al metabolismo aeróbico que utiliza solamente una pequeña proporción del oxígeno contenido en el aire que respiramos. El O2 ocupa el 20% del volumen del aire atmosférico, pero solamente 1 por cada 24 litros del aire inspirado que llega a los pulmones es utilizado para transformar la energía química que se almacena en los tejidos musculares en la energía mecánica que contrae los músculos y hace posible el movimiento del cuerpo humano. Para que tal cosa ocurra el ritmo al que el oxígeno llega a las fibras musculares, a través de la sangre, debe coincidir con el caudal que aquéllas necesitan para poder contraerse. Y esto sólo puede suceder cuando los esfuerzos son de moderada intensidad.
Los glúcidos almacenados en el cuerpo humano permiten un mecanismo alternativo anaeróbico, que no utiliza el oxígeno que proviene del flujo sanguíneo, para generar la energía que necesitan los esfuerzos puntuales, e intensos y para añadir también energía a la que más moderadamente se puede obtener de los metabolismos aeróbicos de los propios glúcidos, en mucha mayor medida de los lípidos y, en proporciones menores, de las proteínas, incrementándose así la potencia total que es posible alcanzar.
En todo caso, provenga la energía de los metabolismos aeróbicos de glúcidos, lípidos o proteínas o de los anaeróbicos alternativos que son exclusivos de los hidratos de carbono, la única sustancia química que provoca realmente el acortamiento de las fibras musculares es el adenosintrifosfato o ATP. Las fibras musculares almacenan pequeñas cantidades de dicha sustancia, que se pueden utilizar instantáneamente, al no necesitar el oxígeno externo transportado, y que son suficientes para producir esfuerzos de muy corta duración, unos 2 segundos, y de muy alta intensidad. Las fibras musculares almacenan también pequeñas cantidades de fosfocreatina, PCr, sustancia que puede ser rápidamente metabolizada para formar ATP y permite ampliar en otros 10 segundos aproximadamente el periodo de intensa actividad muscular anaeróbica.
Los músculos corporales deben regenerar constantemente las reservas agotadas de ATP, porque en otro caso no podrían contraerse y se tomarían rígidos: es el «rigor mortis» que se aprecia en un cuerpo sin vida. La cantidad total de ATP de la que dispone el cuerpo humano es muy pequeña. Pero la cantidad que necesita para sus actividades musculares diarias es muchísimo mayor. Una persona con actividades sedentarias necesita renovar sus reservas de ATP centenares de veces al día.
La mayor parte del oxígeno que almacena un músculo, y que le permite realizar actividades anaeróbicas, se encuentra en la mioglobina, proteína de la familia de la hemoglobina, especializada en almacenar oxígeno en las fibras musculares. Si la carne de pollo, y la de otras aves de corral, es blanca es consecuencia de su sedentarismo: se esfuerzan poco, y no necesitan mioglobina para almacenar el oxígeno. El color oscuro de la carne de la perdiz o del faisán, del jabalí, del venado, y de la caza en general, es debido por el contrario a su «talante nómada» que les exige una intensa actividad muscular, lo que a su vez explica la presencia abundante de mioglobina en sus músculos. Los conejos se mueven por espacios próximos a su madriguera y por ello su carne es más blanca que la de la liebre que utiliza territorios más abiertos y necesita más energía para desplazarse.
Cuando la movilidad de los músculos demanda más oxígeno que el directamente obtenido a través de los pulmones, toma «prestado» el almacenado en las mioglobinas de las fibras musculares, para devolverlo una vez concluido el ejercido. Al hacerlo contrae una «deuda de oxígeno». El atleta que jadea fuertemente al final de un ejercicio intenso, está comenzando a saldar así la deuda contraída, para lo que puede necesitar más o menos tiempo, porque según la intensidad del ejercicio, así será de importante la deuda que tendrá que pagar.
Profundicemos ahora, un poco más acerca de la contribución energética de los tres grupos de nutrientes que incorporamos al alimentarnos a nuestro organismo. Los lípidos o grasas corporales, son el mayor almacén de energía de que dispone el cuerpo humano. Son fundamentalmente triglicéridos, (una molécula de glicerol unida a tres moléculas de ácidos grasos), que para desplazarse por el fluido sanguíneo se unen a determinadas proteínas formando compuestos que son solubles en la sangre. Las grasas sólo se pueden utilizar aeróbicamente y, en general, como combustible de baja intensidad. Suministra la mayor parte de la energía que necesita el cuerpo cuando descansa o realiza actividades de baja o media intensidad. Necesitan un tiempo considerable para acceder a los músculos que los asimilan con relativa lentitud. Sin embargo, como el cuerpo humano almacena entre 50.000 y 200.000 kilocalorías de grasa —unos 135.000 como media—, puede proporcionar energía para cientos de horas de ejercido continuado. No obstante, a pesar de su abundancia, los lípidos no cubren la demanda energética durante actividades musculares de gran intensidad. Y, de hecho, cuando las reservas de hidratos de carbono se han agotado, la oxidación de lípidos sólo aportan la energía necesaria para actividades asociadas al 50% del VO2,max.
Las proteínas están constituidas por largos polímeros de aminoácidos. Si hacemos abstracción del agua, son la fracción celular más importante del organismo. Algunas tienen propiedades catalíticas y actúan como enzimas; otras pueden servir para funciones estructurales, como receptores de señales o para transportar sustancias específicas hacia el interior o el exterior de las células. Son las biomoléculas más versátiles. Pueden contribuir, asimismo, a la generación de energía. Para ello, tienen que eliminar primero el grupo amino. Esto ocurre en el hígado. El carbono «desaminado», que no desanimado, puede convertirse en glucosa en el proceso denominado «gluconeogénesis».
Los glúcidos o hidratos de carbono pueden adoptar la forma más simple de los monosacáridos, azúcar o glucosa, constituidos por seis átomos de carbono unidos a seis moléculas de agua [C6 (H2O)6]. O se pueden almacenar como glucógeno en largas cadenas de polisacáridos [C6 (H2O)6]n o en su forma parcialmente metabolizada que es el lactato.
Utilizados aeróbicamente la glucosa y el glucógeno almacenado en el cuerpo podrían proporcionar durante un par de horas la energía que se necesita para regenerar el ATP. Alternativamente, el glucógeno muscular metabolizado anaeróbicamente en un proceso que se conoce como «glicólisis» solamente aprovecha un 7% de su energía potencial y puede ser agotado en escasos minutos. El glucógeno se caracteriza también por su incapacidad para moverse entre fibras bien abastecidas y otras agotadas. La energía que contiene sólo puede ser transportada en forma de lactatos. Por ello, el lactato no es, como a menudo se le considera, el residuo indeseable, por ser origen de los calambres, agujetas y dolores musculares que limitan la intensidad del ejercido de un atleta. El lactato, en la gluconeogénesis que tiene lugar en el hígado, a donde llega a través del flujo sanguíneo, se vuelve a reconvertir en piruvato, que a su vez, se transforma en glucosa reiniciándose el ciclo metabólico. El lactato es, en consecuencia, un indicador muy valioso del comportamiento muscular y los «umbrales lácticos» indican los límites de la capacidad de generación de energía y los anuncia con dolores crecientes que acaban haciéndose intolerables. Con ejercicios de baja o media intensidad la concentración de lactato en la sangre puede ser de 5 milimoles/litro. En corredores de élite, y en carreras de una hora de duración, suele estar por debajo de 8 mM/l. En otras más cortas, de 10 a 15 minutos, pueden llegar a alcanzar los insoportables 15 mM/l.
A modo de síntesis, para concluir este apartado, podemos reflejar, en el cuadro siguiente y en grandes trazos, los procesos metabólicos aeróbicos que permiten la producción de ATP, a partir de los glúcidos, lípidos y proteínas que ingerimos con los alimentos así como los anaeróbicos, con los que, alternativamente, los glúcidos pueden regenerar más rápidamente las reservas de ATP.
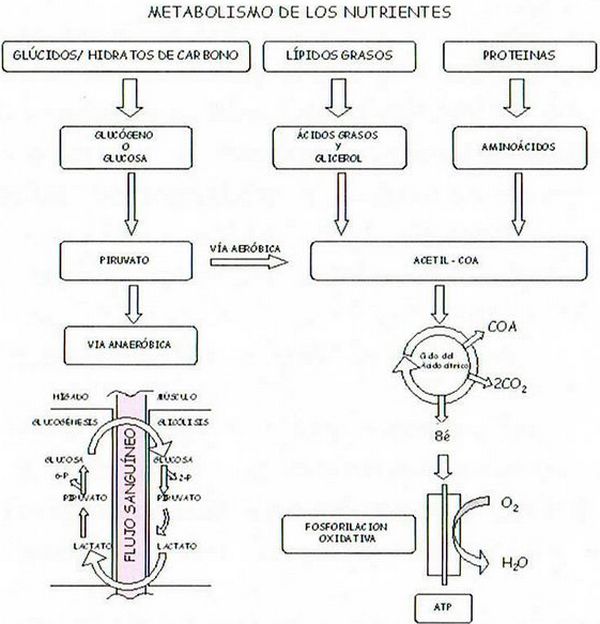
En los procesos de digestión, las grandes moléculas de los glúcidos, se dividen en moléculas de glucosa u otros monosacáridos; los lípidos se hidrolizan transformándose en ácidos grasos y glicerol; las proteínas, se hidrolizan también en sus 20 clases de aminoácidos constituyentes. A continuación, la mayoría de estas moléculas se convierten en el fragmento acetilo del acetil-CoA, cuya oxidación completa se produce primero en el ciclo del ácido nítrico, en el que se desprende la coenzima A y dos moléculas de CO2 y después en la fosforilación oxidativa que concluye con la producción de ATP.
En el caso específico de la glucosa, por un complejo proceso que engloba diez reacciones químicas y que se denomina glicólisis, se producen piruvatos que, por vía aeróbica, se transforman en las moléculas de acetil-CoA. Cuando el aporte de oxígeno no es suficiente la glucosa se metaboliza anaeróbicamente en moléculas de lactato, que a su vez llegan por el fluido sanguíneo al hígado donde, en el proceso denominado «gluconeogénesis», se reconvierte en piruvato y, a continuación, en glucosa iniciándose de nuevo el ciclo que acaba con la regeneración de ATP que provoca la contracción de los músculos. Los valores medios de la energía almacenada (en kilocalorías) por los diferentes nutrientes en los músculos, en los tejidos adiposos y en el hígado de un adulto de unos 70 kg de masa corporal son los que se muestran en el cuadro siguiente:
|
Glucosa o Glucógeno |
Triglicéridos |
Proteínas movilizables |
|
|---|---|---|---|
|
Músculos |
1.200 |
450 |
24.000 |
|
Tejidos adiposos |
80 |
13.500 |
40 |
|
Hígado |
400 |
450 |
400 |
Por otra parte, la velocidad máxima de producción de ATP, en milimoles[2] por segundo para una persona también de 70 kg con una masa muscular del 40%, son:
|
Fuente de energía |
Velocidad de producción de ATP (mmol/s) |
|---|---|
|
PCr |
73,3 |
|
Conversión del glucógeno muscular en lactato |
39,1 |
|
Conversión del glucógeno muscular en CO2 |
16,7 |
|
Conversión del glucógeno hepático en CO2 |
6,2 |
|
Conversión de los ácidos grasos del tejido adiposo en CO2 |
6,7 |
El ritmo de regeneración de las moléculas de ATP es más de 10 veces superior cuando el organismo utiliza la fosfocreatina, PCr, que cuando recurre a la conversión de las grasas en CO2 en los parsimoniosos catabolismos aeróbicos.