Capítulo 8
El darwinista, aunque educado, es a lo sumo un mono afeitado.
WILLIAM S. GILBERT Y ARTHUR SULLIVAN, Princess Ida
En 1924, mientras se vestía para una boda, Raymond Dart recibió lo que habría de ser el mayor descubrimiento fósil del siglo XX. Dart era un joven profesor de anatomía en la Universidad de Witwatersrand, en Sudáfrica, pero también un antropólogo aficionado que había hecho correr la voz de que estaba buscando «hallazgos interesantes» para llenar un nuevo museo de anatomía. Mientras se vestía el esmoquin, el cartero le trajo dos cajas de rocas que contenían fragmentos de huesos excavados en una cantera de caliza cercana a Taungs, en la región del Transvaal. En sus memorias, Adventures with the Missing Link, Dart describe aquel momento:
Tan pronto como levanté la tapa, un escalofrío de excitación me recorrió el cuerpo. En la parte superior de la roca había lo que sin duda era el molde del interior de un cráneo. Si hubiera sido el molde endocraneal fosilizado de cualquier especie de simio, hubiera sido un gran descubrimiento, pues nunca antes se había hallado tal cosa. Pero supe desde el primer momento que lo que tenía en mis manos no era un cerebro antropoide normal y corriente. Lo que tenía, en arena consolidada con caliza, era la réplica de un cerebro tres veces más grande que el de un babuino y considerablemente mayor que el de un chimpancé adulto. Podía apreciarse con claridad la singular imagen de las circunvoluciones y surcos del cerebro y los vasos sanguíneos del cráneo.
No era lo bastante grande para un hombre primitivo, pero incluso para un simio era un cerebro grande y abultado y, lo que era más importante, el prosencéfalo era tan grande y había crecido tanto hacia atrás que cubría casi completamente el rombencéfalo.
¿Había allí, en aquel montón de rocas, alguna cara que correspondiera al cerebro? Rebusqué febrilmente en las cajas. Mi afán se vio recompensado, pues encontré una gran piedra con una depresión en la que el molde encajaba a la perfección. Vagamente visible en la piedra había el perfil de un trozo del cráneo e incluso la parte posterior de la mandíbula inferior y un alvéolo de un diente que me decía que el rostro debía hallarse por algún lado en aquel bloque…
Permanecí en la sombra sosteniendo el cerebro con la misma codicia con la que un avaro abraza su oro, mientras mi mente se disparaba. Tenía, de eso estaba seguro, uno de los hallazgos más importantes jamás realizados en la historia de la antropología.
La desacreditada teoría de Darwin de que los antiguos progenitores del hombre probablemente habían vivido en África me vino a la mente. ¿Iba a ser yo el instrumento del hallazgo de su «eslabón perdido»?
Estas agradables ensoñaciones fueron interrumpidas por el novio, que me tiraba de la manga.
«Por Dios, Ray», me dijo, intentando reprimir la urgencia nerviosa de su voz. «Tienes que acabar de vestirte inmediatamente, o tendré que buscarme otro padrino. El coche nupcial llegará en cualquier momento.»
La preocupación del novio es comprensible. Nadie quiere descubrir el día de su boda que su padrino está más interesado en una caja llena de piedras polvorientas que en su inminente boda. Pero es difícil no empatizar también con Dart. En El origen del hombre, Darwin había conjeturado que nuestra especie se había originado en África porque nuestros parientes más próximos, los gorilas y los chimpancés, se encontraban allí. Pero era poco más que un presentimiento. No había fósiles que lo respaldaran. Y entre nosotros y el antepasado común que debíamos haber compartido con los grandes simios, un ancestro con más aspecto de simio que de humano, se abría una manifiesta brecha. Aquel día de 1924 salió a la luz la primera piedra del puente que con el tiempo nos permitiría cruzar el abismo: allí, en las temblorosas manos de Dart, podía verse directamente lo que hasta entonces se había conocido de manera simplista como «el eslabón perdido». Hay que preguntarse cómo pudo atender a sus obligaciones durante la ceremonia.
Lo que Dart había encontrado en aquella caja era el primer espécimen de lo que más tarde se llamaría Australopithecus africanus («hombre simio austral»). Durante los tres meses siguientes, Dart realizó una meticulosa disección de la roca con la ayuda de unas agujas de calceta afiladas que le tomó prestadas a su mujer, y consiguió revelar el rostro completo. Era la cara de un niño, hoy conocido como «niño de Taung», en el que podían verse incluso los dientes de leche y los molares que comenzaban a salir. Su mezcla de caracteres de humano y de simio confirmaron la idea de Dart de que efectivamente había tropezado con los albores de la ascendencia humana.
Desde los tiempos de Dart, paleoantropólogos, genetistas y biólogos moleculares han utilizado fósiles y secuencias de ADN para determinar nuestro lugar en el árbol de la evolución. Somos simios que descendemos de otros simios, y nuestro primo más cercano es el chimpancé, cuyos antepasados divergieron de los nuestros hace varios millones de años en África. Éstos son hechos indisputables que, lejos de disminuir nuestra humanidad, deberían producir satisfacción y admiración, pues nos conectan con todos los organismos, los vivos y los muertos.
Pero no todo el mundo lo ve de esta manera. Para quienes se muestran reacios a aceptar el darwinismo, la evolución humana constituye el núcleo de su resistencia. No parece tan difícil aceptar que los mamíferos evolucionaron a partir de los reptiles, o los animales terrestres a partir de los peces. Pero nos cuesta llegar a aceptar que, igual que cualquier otra especie, evolucionamos a partir de un antepasado que era muy distinto. Siempre nos hemos visto como si, de algún modo, estuviéramos al margen del resto de la naturaleza. Animados por la creencia religiosa de que los seres humanos son objeto especial de la creación, y por un solipsismo natural que acompaña a un cerebro que es consciente de sí mismo, nos resistimos a aceptar la lección evolutiva de que, como el resto de los animales, somos el producto contingente del proceso ciego y mecánico de un proceso de selección natural. Y a causa de la hegemonía de la religión fundamentalista en Estados Unidos, este país se encuentra entre los que más se resisten a la evidencia de la evolución humana.
En el famoso «Juicio del Mono» de 1925, el profesor de enseñanza secundaria John Scopes fue juzgado en Dayton (Tennessee), y hallado culpable de violar la Ley Butler de este estado. Curiosamente, esta ley no prohibía enseñar la evolución en general, sino sólo la idea de que los humanos habían evolucionado:
Queda promulgado por la Asamblea General del Estado de Tennessee que será ilegal que cualquier profesor de cualquiera de las universidades, escuelas normales y cualesquiera otras escuelas públicas del Estado que estén financiadas en todo o en parte por los fondos para la escuela pública del Estado, enseñe cualquier teoría que niegue la historia de la Divina Creación del hombre tal como la enseña la Biblia, y enseñe en su lugar que el hombre desciende de un orden inferior de animales.
Aunque los creacionistas más liberales admiten que algunas especies podrían haber evolucionado a partir de otras, todos los creacionistas trazan la divisoria en los humanos. La brecha entre nosotros y otros primates, dicen, no puede salvarla la evolución: por necesidad ha intervenido un acto de creación especial.
La idea de que los humanos forman parte de la naturaleza ha sido anatema durante la mayor parte de la historia de la biología. En 1735 el botánico sueco Carolus Linnaeus, que estableció el sistema de clasificación biológica, agrupó a los humanos, que llamó Homo sapiens («hombre sabio»), con los monos y los simios sobre la base de sus semejanzas anatómicas. Linnaeus no sugirió una relación evolutiva entre estas especies, pues su intención explícita era revelar el orden subyacente a la creación de Dios; aun así, su decisión fue controvertida, y provocó la ira de su arzobispo.
Un siglo más tarde, Darwin sabía bien que despertaría la ira al sugerir, como firmemente creía, que los humanos habían evolucionado a partir de otras especies. En El origen ventiló la cuestión con mucho tiento, colando al final del libro una sentencia oblicua: «Se arrojará luz sobre el origen del hombre y su historia». Darwin no abordó de pleno la cuestión hasta más de una década más tarde en El origen del hombre (1871). Envalentonado por su creciente perspicacia y convicción, y por la confianza que había ganado gracias a la rápida aceptación de sus ideas, por fin hizo públicas sus ideas de manera explícita. Tras recopilar pruebas y observaciones de la anatomía y el comportamiento, Darwin afirmó no sólo que los humanos habían evolucionado a partir de organismos con aspecto de simios, sino que lo habían hecho en África:
Llegamos así a la conclusión de que los humanos descienden de un cuadrúpedo peludo, dotado de una cola y con las orejas puntiagudas, probablemente de hábitos arborícolas, y que habitaba en el Viejo Mundo.
Con qué fuerza debió golpear esa frase los oídos Victorianos. ¡Pensar que nuestros antepasados vivían en los árboles! ¡Y que estaban dotados de una cola y orejas puntiagudas! En su último capítulo, Darwin se enfrenta por fin a las objeciones religiosas:
Soy consciente de que las conclusiones a las que se llega en esta obra serán denunciadas por algunos por ser muy irreligiosas; pero quien las denuncie deberá mostrar por qué es más irreligioso explicar el origen del hombre como especie distinta a partir de alguna forma inferior, por medio de las leyes de la variación y la selección natural, que explicar el nacimiento del individuo por medio de las leyes ordinarias de la reproducción [las pautas del desarrollo].
No obstante, nunca logró convencer a todos sus colegas. Alfred Russel Wallace y Charles Lyell, competidor y mentor de Darwin, respectivamente, se apuntaron a la idea de la evolución pero no quedaron convencidos de que la selección natural pudiera explicar las facultades mentales superiores de los humanos. Hicieron falta fósiles para convencer por fin a los escépticos de que los humanos realmente habían evolucionado.
Ancestros fósiles
En 1871, el registro fósil humano comprendía únicamente unos pocos huesos de neandertales, de aparición reciente y con aspecto demasiado humano para que sirvieran de eslabón perdido entre nosotros y los simios. De hecho, fueron interpretados como una población aberrante de Homo sapiens. En 1891, el médico holandés Eugène Dubois descubrió en Java un casquete craneal, algunos dientes y un hueso de cadera que se ajustaban a lo esperado: el cráneo era algo más robusto que el de los humanos modernos, y el cerebro de tamaño menor. Pero nervioso por la oposición religiosa y científica a sus ideas, Dubois volvió a enterrar los huesos de Pithecanthropus erectus (hoy Homo erectus) bajo su casa, ocultándolos a la curiosidad científica durante tres décadas.
El descubrimiento por Dart del niño de Taung desató una caza de ancestros humanos en África que culminó en las célebres excavaciones de los Leakey en la garganta de Olduvai a principios de la década de 1930, el descubrimiento de «Lucy» por Donald Johanson en 1974, y varios otros hallazgos. Hoy disponemos de un registro fósil razonablemente bueno de nuestra evolución, aunque está lejos de ser completo. Hay en él, como veremos, muchos misterios, y más de unas pocas sorpresas.
Pero incluso sin los fósiles sabríamos algo sobre nuestro lugar en el árbol de la evolución. Tal como propuso Linnaeus, nuestra anatomía nos sitúa en el orden Primates junto a los monos, los simios y los lémures, todos los cuales comparten rasgos como los ojos en posición frontal, las uñas, la visión en color y los pulgares oponibles. Otras características nos sitúan en la superfamilia Hominoideos junto a los «simios menores» (gibones) y los «grandes simios» (chimpancés, gorilas, orangutanes y nosotros mismos). Y dentro de los Hominoideos, quedamos agrupados con los grandes simios en la familia Homínidos, que comparten características únicas como las uñas planas, treinta y dos dientes, ovarios engrosados y cuidados parentales prolongados. Estas características compartidas muestran que nuestro antepasado común con los grandes simios vivió más recientemente que nuestro antepasado común con cualquier otro mamífero.
Los datos moleculares derivados de la secuenciación del ADN y de proteínas confirman estas relaciones, y nos dicen también de manera aproximada cuándo divergimos de nuestros parientes. Nuestro pariente más próximo es el chimpancé (igual distancia del común que del bonobo) y divergimos de nuestro antepasado común hace unos 7 millones de años. El gorila es un pariente ligeramente más lejano, y los orangutanes todavía más (12 millones de años desde el antepasado común).
Para muchos, sin embargo, las pruebas fósiles son psicológicamente más convincentes que los datos moleculares. Una cosa es enterarse de que compartimos un 98,5 por 100 de nuestro ADN con los chimpancés y otra completamente distinta ver el esqueleto de un australopitecino, con su pequeño cráneo con aspecto de simio encima de un esqueleto casi idéntico al de los humanos modernos. Pero antes de mirar los fósiles podemos hacer algunas predicciones sobre lo que cabe encontrar en ellos si los humanos han evolucionado a partir de los simios.
¿Qué aspecto debería tener nuestro «eslabón perdido»? Recuérdese que el «eslabón perdido» es la especie ancestral singular que dio origen a los humanos modernos, de un lado, y a los chimpancés, del otro. No es razonable esperar descubrir justamente esa especie crucial, pues su identificación requeriría una serie completa de fósiles de ancestros y descendientes tanto en el linaje de los chimpancés como en el humano, unas series que entonces podríamos seguir en el pasado hasta el punto donde se crucen. Salvo por unos pocos microorganismos marinos, no existen secuencias de fósiles tan completas. Además, nuestros antepasados humanos más antiguos eran grandes, vivían en densidades bajas en comparación con los herbívoros como los antílopes, y habitaban en una pequeña parte de África en unas condiciones secas que no son propicias para la fosilización. Sus fósiles, como los de todos los simios y monos, son escasos. Esto recuerda el problema de la evolución de las aves, con formas transicionales también poco frecuentes. Aunque podemos seguir la evolución de las aves desde los reptiles con plumas, no estamos seguros de qué especies fósiles concretas fueron antepasados directos de las aves modernas.
A la vista de todo esto, no cabe esperar encontrar la especie exacta que representa el «eslabón perdido» entre los humanos y otros simios. Sólo podemos esperar encontrar sus primos evolutivos. Hay que recordar también que ese antepasado común no era un chimpancé, y probablemente no tenía el aspecto ni de los chimpancés modernos ni de los humanos actuales. Con todo, es probable que el «eslabón perdido» tuviera un aspecto más cercano al de los chimpancés modernos que al de los humanos modernos. Somos un poco el bicho raro de la evolución de los simios modernos, pues todos se parecen más entre sí de lo que se parecen a nosotros. Los gorilas son nuestros primos distantes, y aun así comparten con los chimpancés características como el cerebro relativamente pequeño, el pelo en todo el cuerpo, el caminar sobre los nudillos, y los caninos grandes y afilados. Los gorilas y los chimpancés también tienen una «arcada dentaria rectangular»: vista desde arriba, la fila inferior de sus dientes recuerda los tres lados de un rectángulo (véase la Figura 27). Los humanos son la especie que ha divergido del plan corporal de los simios: tenemos pulgares flexibles, muy poco pelo, caninos más pequeños y romos, y caminamos erectos. Nuestra hilera de dientes no es rectangular, sino parabólica, como cualquiera puede ver si se mira los dientes en el espejo. Lo más llamativo, sin embargo, es que tenemos un cerebro mucho más grande que el de cualquier simio: el cerebro del chimpancé adulto tiene un volumen de unos 450 centímetros cúbicos, el de un humano moderno, unos 1.450 centímetros cúbicos. Cuando comparamos las similitudes de los chimpancés, gorilas y orangutanes y los caracteres divergentes de los humanos, podemos concluir que, respecto a nuestro antepasado común, hemos cambiado más que los simios modernos.
Así pues, cabe esperar encontrar hace alrededor de 5 a 7 millones de años antepasados fósiles con caracteres compartidos por los chimpancés, orangutanes y gorilas (estos caracteres son compartidos porque estaban presentes en el antepasado común), pero con algunas características humanas. A medida que los fósiles sean más recientes, deberíamos ver cómo los cerebros se hacen relativamente mayores, los caninos menores, la hilera de dientes menos rectangular y más curva, y la postura más erecta. Y eso es exactamente lo que vemos. Aunque lejos de estar completo, el registro de la evolución humana es una de las mejores confirmaciones que tenemos de una predicción evolutiva, y es especialmente gratificante porque la predicción viene del propio Darwin.
Pero primero algunas advertencias. No disponemos (ni esperamos disponer) de un registro fósil continuo de la ascendencia de los humanos. Lo que vemos en su lugar es un enmarañado arbusto con muchas especies distintas. La mayoría de ellas se extinguieron sin dejar especies descendientes, y sólo un linaje genético siguió su camino en el tiempo hasta convertirse en los humanos modernos. Todavía no estamos seguros de qué especies fósiles se sitúan a lo largo de ese linaje concreto, y cuáles, en cambio, fueron callejones sin salida de la evolución. Lo más sorprendente que hemos aprendido sobre nuestra historia es que hemos tenido muchos primos evolutivos que se extinguieron sin dejar descendientes. Es posible incluso que hasta cuatro especies humanoides habitaran en África al mismo tiempo, y quizá en el mismo lugar. ¡Qué encuentros se pueden haber producido! ¿Se habrán intentado matar unos a otros, o habrán intentado cruzarse?
Los nombres de los fósiles humanos ancestrales tampoco pueden tomarse demasiado en serio. Como la teología, la paleoantropología es una disciplina en la que el número de estudiosos supera en mucho el de objetos de estudio. Abundan los debates acalorados, a veces enconados, sobre si un determinado fósil es realmente algo nuevo o sólo una simple variante de una especie ya nombrada. Estas disputas sobre los nombres científicos suelen revestir poca importancia. Que un fósil humanoide sea clasificado como una especie u otra puede depender de cuestiones tan minúsculas como medio milímetro del diámetro de un diente, o ligeras diferencias en la forma de la pelvis. El problema es, sencillamente, que hay demasiados pocos especímenes distribuidos por un área geográfica demasiado grande como para poder tomar decisiones fiables. Constantemente se producen hallazgos nuevos o se revisan conclusiones anteriores. Lo que no debemos perder de vista es la tendencia general de los fósiles a lo largo del tiempo, que muestra claramente un cambio desde caracteres simiescos a caracteres humanos.
Vayamos a por los huesos. Los antropólogos aplican el término homínino a todas las especies del lado «humano» de nuestro árbol genealógico después de la separación de la rama que condujo a los chimpancés modernos.[46] En la actualidad se reconocen como especies veinte tipos de homíninos; quince de ellos se presentan en la Figura 24 en el orden aproximado en que aparecieron. La Figura 25 muestra los cráneos de unos pocos homíninos representativos, junto al de los chimpancés modernos y los humanos, como referencia.
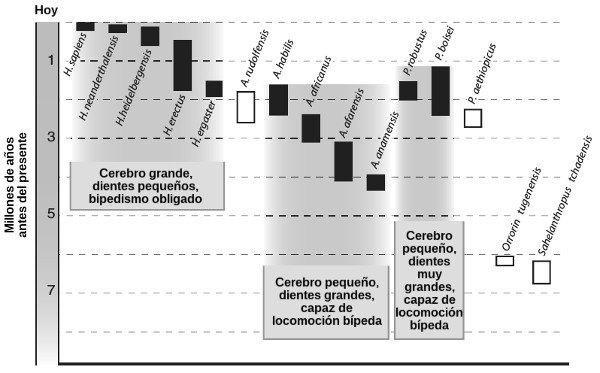
FIGURA 24. Quince especies de homíninos, los periodos en los que aparecen como fósiles, y la naturaleza de su cerebro, dientes y locomoción. Los fósiles representados con cajas vacías son demasiado fragmentarios como para extraer conclusiones sobre su locomoción y volumen cerebral.
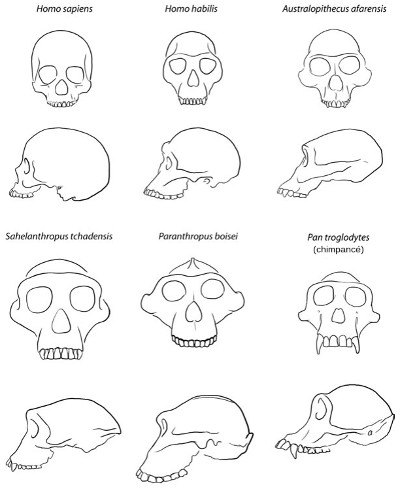
FIGURA 25. Cráneos de humanos modernos (Homo sapiens), homíninos fósiles y un chimpancé (Pan troglodytes).
Nuestra pregunta principal es, por supuesto, cómo determinar la pauta de la evolución humana. ¿Cuándo encontramos los fósiles más antiguos que pudieran representar a nuestros antepasados cuando ya habían divergido de otros simios? ¿Cuáles de nuestros parientes homíninos se extinguieron, y cuáles fueron nuestros antepasados directos? ¿Cómo se convirtieron los rasgos del simio ancestral en los de los humanos actuales? ¿Evolucionó primero nuestro voluminoso cerebro o nuestra postura erguida? Sabemos que los humanos comenzaron a evolucionar en África, pero ¿qué parte de nuestra evolución se produjo en otros lugares?
Con la salvedad de algunos fragmentos de huesos cuya clasificación no está clara, hasta hace poco el registro fósil de los homíninos no se remontaba más allá de hace 4 millones de años. Pero en 2002, Michel Brunet y sus colaboradores anunciaron el sorprendente descubrimiento de un posible homínino más antiguo, Sahelanthropus tchadensis, de los desiertos del Chad, en el África central, en la región conocida como Sahel. Lo más sorprendente de este hallazgo es su datación: hace de 6 a 7 millones de años, justo cuando las pruebas moleculares nos dicen que nuestro linaje se separó del de los chimpancés. Sahelanthropus podría representar el antepasado humano más antiguo, pero también podría ser una rama lateral que se extinguió. Su mezcla de caracteres, sin embargo, parece situarlo claramente en el lado humano de la divisoria con los chimpancés. Lo que tenemos en este caso es un cráneo casi completo (aunque un poco aplastado por la fosilización) que es un mosaico, una curiosa mezcla de caracteres de simios y de humanos. Como los simios, el cráneo es alargado y el cerebro pequeño, como el del chimpancé; pero igual que los homíninos posteriores, tiene el rostro plano, los dientes pequeños y las cejas prominentes (Figura 25).
Puesto que no tenemos el resto del esqueleto, no podemos decir si Sahelanthropus poseía la crucial habilidad de caminar erecto, pero hay una interesante indicación de que podría ser así. En los simios que caminan apoyándose en los nudillos, como los gorilas y los chimpancés, la postura habitual del animal es horizontal, de manera que la médula espinal penetra en el cráneo por detrás. En los humanos erectos, sin embargo, el cráneo se asienta directamente encima de la médula espinal. Puede apreciarse esta diferencia en la posición de la abertura del cráneo por la que pasa la médula espinal (el foramen magnum, o «agujero grande», en latín): este orificio está en posición más frontal en los humanos. En Sahelanthropus, el orificio es más frontal que en los simios que caminan apoyándose en los nudillos. Esto es muy interesante, porque si esta especie realmente se encuentra en el lado homínino de la divisoria, nos sugiere que la locomoción bípeda fue una de las primeras innovaciones evolutivas que nos distinguieron de los otros simios.[47]
Después de Sahelanthropus, tenemos unos pocos fragmentos de hace unos 6 millones de años de otra especie, Orrorin tugenensis, que comprenden un solo hueso de la pierna que se ha interpretado como prueba de bipedismo. Pero luego se abre una brecha de 2 millones de años sin ningún fósil homínino sustantivo. Es en este período donde algún día encontraremos información crucial sobre cuándo comenzamos a caminar erguidos. Pero hace 4 millones de años reaparecen los fósiles, y vemos cómo comienza a ramificarse el árbol de los homíninos. De hecho, podrían haber vivido al mismo tiempo varias especies. Entre ellas están los australopitecinos gráciles, que una vez más presentan una mezcla de caracteres simiescos y humanoides. Del lado de los simios tienen el cerebro de tamaño parecido al de un chimpancé, y su cráneo es más parecido al de un simio que al de un humano. Pero los dientes son relativamente pequeños, y se disponen en hileras a medio camino entre la forma rectangular de los simios y el paladar parabólico de los humanos. Y sin lugar a dudas eran bípedos.
Un conjunto de fósiles antiguos de Kenia, agrupados bajo la denominación de Australopithecus anamensis, muestra interesantes indicaciones de bipedismo en un único fémur fosilizado. Pero el hallazgo definitivo fue realizado por el paleontólogo norteamericano Donald Johanson mientras hacía prospección de fósiles en la región de Afar, en Etiopía. La mañana del 30 de noviembre de 1974, Johanson se levantó sintiéndose afortunado, y tomó nota de ello en su diario de campaña. Pero no podía ni imaginar la suerte que tendría. Tras buscar en vano durante toda la mañana en un barranco seco, Johanson y Tom Gray, un estudiante de doctorado, estaban a punto de dejarlo por aquel día y regresar al campamento. De repente, Johanson vio un hueso de homínido en el suelo, y luego otro, y otro más. Para su asombro, habían tropezado con los huesos de un solo individuo, que más tarde recibió la denominación formal de AL 288-1, pero que se conoce de manera más familiar como «Lucy», en honor a la canción de los Beatles «Lucy in the Sky with Diamonds», que se escuchó repetidas veces en el campamento durante la celebración del hallazgo.
Cuando se montaron los centenares de fragmentos de Lucy, resultó ser una hembra de una nueva especie, Australopithecus afarensis, que databa de hace 3,2 millones de años. Tenía entre veinte y treinta años de edad, medía poco más de un metro, pesaba apenas treinta kilos y posiblemente sufría artritis. Pero lo más importante es que caminaba sobre dos piernas.
¿Cómo lo sabemos? Por la manera en que el fémur se une a la pelvis por uno de sus extremos y a la rodilla por el otro (Figura 26). En un primate bípedo como nosotros, los fémures forman un ángulo hacia el interior, acercándose desde la pelvis, de manera que el centro de gravedad se mantiene en un mismo lugar mientras se camina, lo que permite un eficiente paso bípedo adelante y atrás. En los simios que caminan apoyándose en los nudillos, los fémures se abren ligeramente, dándoles un aspecto patizambo. Cuando intentan caminar erguidos, se tambalean torpemente, como el pequeño vagabundo de Charlie Chaplin.[48] Así pues, si cogemos un fósil de primate y nos fijamos en cómo se une el fémur a la pelvis, podremos decir si caminaba sobre dos patas o sobre cuatro. Si los fémures se inclinan hacia el centro, era bípedo. Así pasa en Lucy, y casi con el mismo ángulo que en los humanos modernos. Caminaba erguida. También su pelvis se parece mucho más a la de los humanos modernos que a la de los chimpancés.
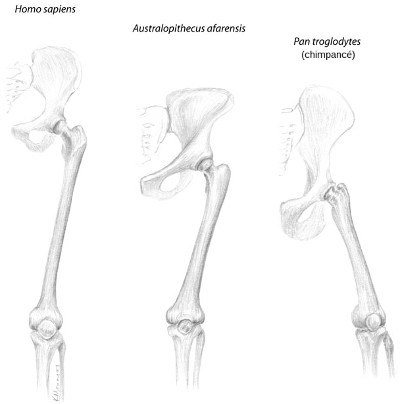
FIGURA 26. Unión del fémur (el hueso largo de la pierna) con la pelvis en los humanos modernos, los chimpancés y Australopithecus afarensis. La pelvis de A. afarensis es intermedia entre las otras dos, pero su fémur inclinado hacia dentro (una indicación de que caminaba erecto) se parece al de los humanos y contrasta con el fémur abierto del chimpancé, que camina apoyándose en los nudillos.
Un equipo de paleoantropólogos dirigido por Mary Leakey confirmó el bipedismo de A. afarensis con otro hallazgo notable realizado en Tanzania: las famosas «huellas de Laetoli». En 1976, Andrew Hill y otro miembro del equipo estaban disfrutando de un descanso dedicándose al pasatiempo favorito del campamento: lanzarse unos a otros cachos de boñiga seca de elefante. Mientras buscaba munición en un lecho de río seco, Hill tropezó con una hilera de huellas fosilizadas. Tras una cuidadosa excavación, las huellas resultaron ser parte de un rastro de unos veinticinco metros que habían dejado un par de homíninos que claramente habían estado caminando sobre dos piernas (no había impresiones de nudillos) durante una tormenta de ceniza causada por la erupción de un volcán. Aquella tormenta fue seguida por la lluvia, que convirtió la ceniza en una capa parecida al cemento que más tarde quedó sellada por otra capa de ceniza seca, preservando las huellas.
Las huellas de Laetoli son virtualmente idénticas a las que hace un humano moderno al caminar sobre un suelo blando. Y los pies eran casi con certeza de unos parientes de Lucy: las huellas tienen la medida correcta y el rastro se remonta a hace 3,6 millones de años, cuando el único homínino conocido era A. afarensis. Lo que tenemos aquí es el más raro de los hallazgos: comportamiento humano fosilizado.[49] Uno de los rastros es de huellas mayores que el otro, por lo que deben provenir de un macho y una hembra (sabemos por otros fósiles de afarensis que presentaban dimorfismo sexual en la talla). Las huellas de la hembra parecen un poco más profundas en un lado que en el otro, lo que quizá indique que llevaba un niño en la cadera. El rastro evoca la imagen de una pareja pequeña y peluda que cruza el llano durante una erupción volcánica. ¿Estaban asustados y se cogían de la mano?
Al igual que otros australopitecinos, Lucy tenía la cabeza simiesca, con un volumen craneal como el de un chimpancé. Pero su cráneo muestra también algunas trazas más humanas, por ejemplo una arcada dentaria semiparabólica y caninos de tamaño más reducido (Figuras 25 y 27). Entre la cabeza y la pelvis tenía una mezcla de caracteres propios de simios y de humanos: los brazos eran, en términos relativos, más largos que los de los humanos modernos, pero más cortos que los de los chimpancés, y las falanges de los dedos eran un tanto curvadas, como las de los simios. Esto ha llevado a suponer que afarensis debía pasar al menos una parte de su tiempo en los árboles.
Uno no podría pedir una forma transicional mejor entre los humanos y los antiguos simios que Lucy. Del cuello para arriba, es simio; en la parte media, una mezcla; y de la cintura para abajo, casi un humano moderno. Además, nos proporciona un dato crucial sobre su evolución: nuestra postura erecta evolucionó mucho antes que nuestro cerebro grande. En el momento de su descubrimiento, esto iba en contra de la suposición generalmente admitida de que el cerebro grande había evolucionado primero, y nos hizo repensar el modo en que la selección natural puede haber modelado a los humanos modernos.
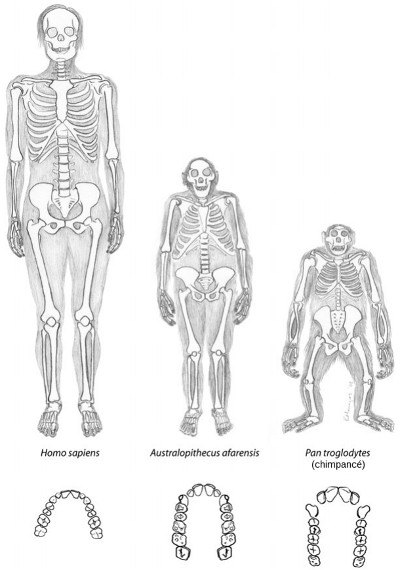
FIGURA 27. Esqueletos y arcadas dentarias del moderno Homo sapiens, Australopithecus afarensis («Lucy») y un chimpancé. Aunque los chimpancés no son antepasados del linaje humano, probablemente se parezcan al antepasado común más que los humanos. En muchos sentidos, A. afarensis es intermedio entre la morfología de simio y la de humano.
Después de A. afarensis, el registro fósil muestra una confusa mezcolanza de especies australopitecinas gráciles que se prolonga hasta hace unos 2 millones de años. La secuencia cronológica muestra una progresión hacia una forma humana más moderna: la hilera de dientes se hace más parabólica, el cerebro aumenta de tamaño y el esqueleto pierde sus caracteres simiescos.
Luego todo se vuelve más confuso, pues hace 2 millones de años se encuentra la frontera entre los fósiles situados en el género Australopithecus y los situados en el género más moderno Homo. No debemos pensar, sin embargo, que este cambio de nombres significa que se produjo algún cambio transcendental, que de algún modo los «verdaderos humanos» evolucionaron de un solo golpe. Que un fósil reciba un nombre u otro depende de si tiene un cerebro mayor (Homo) o menor (Australopithecus), por lo general con arreglo a una frontera arbitraria de alrededor de 600 centímetros cúbicos. Algunos fósiles australopitecinos, como A. rudolfensis, tienen un volumen cerebral tan intermedio, que los científicos debaten acaloradamente sobre si debería llamarse Homo o Australopithecus. Este problema de nomenclatura se agrava con el hecho de que incluso dentro de una misma especie vemos una considerable variación en el tamaño del cerebro. (Los humanos modernos, por ejemplo, muestran un amplio abanico de volúmenes, entre 1.000 y 2.000 centímetros cúbicos, que, por cierto, no guardan correlación con la inteligencia.) Pero las dificultades semánticas no deberían distraemos de entender que los últimos australopitecinos, que ya eran bípedos, comenzaban a mostrar cambios en los dientes, el cráneo y el cerebro que presagian a los humanos modernos. Es muy probable que el linaje que dio origen a los humanos modernos incluya al menos una de estas especies.
Otro salto importante en la evolución humana fue la habilidad para fabricar y utilizar instrumentos. Aunque los chimpancés utilizan instrumentos simples, por ejemplo palitos para extraer termitas de sus montículos, el uso de instrumentos más elaborados seguramente requería unos pulgares más flexibles y una postura erecta que dejara las manos libres. El primer humano que sin ninguna duda utilizaba instrumentos fue Homo habilis (Figura 25), cuyos restos aparecen por primera vez hace unos 2,5 millones de años. H. habilis significa, como es obvio, «hombre hábil», y sus fósiles aparecen asociados a diversos instrumentos de piedra o lascas que utilizaban para cortar, raspar y matar a sus presas. No estamos seguros de que esta especie sea un antepasado directo de H. sapiens, pero habilis desde luego presenta cambios hacia una condición más humana, por ejemplo en los molares más reducidos y el cerebro más voluminoso que en los australopitecinos. Un molde de un cerebro muestra unos claros abultamientos que corresponden al área de Broca y el área de Wernicke, las partes del lóbulo izquierdo del cerebro asociados con la producción y comprensión del habla. Estos bultos suscitan la posibilidad, todavía muy lejos de ser cierta, de que habilis fuese la primera especie con un lenguaje hablado.
Lo que sí sabemos es que H. habilis coexistió, si no en el espacio, al menos en el tiempo, con varios otros homíninos. Los más famosos son los homíninos «robustos» (en oposición a los gráciles) del África oriental. Comprenden al menos tres: Paranthropus (o Australopithecus) boisei (Figura 25), P. robustus y P. aethiopicus, todos ellos con grandes cráneos, pesados dientes masticadores (algunos de los molares tenían más de dos centímetros y medio de ancho), huesos recios y cerebro relativamente pequeño. También tenían una cresta sagital: una prominencia del hueso que recorría la parte superior del cráneo y servía para anclar los grandes músculos masticadores. Algunas especies robustas probablemente subsistieron con alimentos duros o correosos, por ejemplo raíces, nueces y tubérculos (P. boisei, descubierto por Louis Leakey, recibió el apelativo de «hombre cascanueces»). Las tres especies se extinguieron hace aproximadamente 1,1 millones de años sin dejar ninguna especie descendiente.
Pero H. habilis quizá conviviera también con tres especies más de Homo: H. ergaster, H. rudolfensis y H. erectus, aunque cada una de estas tres especies muestra una considerable variación y las relaciones entre ellas son discutidas. H. erectus («hombre erguido») ostenta la distinción de ser el primer homínino que salió de África: se han encontrado restos de esta especie en China («hombre de Pequín»), Indonesia («hombre de Java»), Europa y Oriente Medio. Es probable que a medida que sus poblaciones africanas se expandían, erectus simplemente buscara nuevas tierras donde vivir.
Para cuando se produjo esta diáspora, el tamaño del cerebro de erectus era prácticamente igual al de los humanos modernos. Sus esqueletos también eran casi idénticos a los nuestros, aunque todavía tenían el rostro plano, sin barbilla (el mentón es una característica de H. sapiens). Sus utensilios eran complejos, sobre todo los de los últimos erectus, que fabricaban complejas hachas y raspadores de piedra con intricados lascados. La especie también parece ser responsable de uno de los momentos más transcendentales de la historia cultural humana: el control del fuego. En una cueva de Swartkrans, en Sudáfrica, los científicos han hallado restos de erectus junto a huesos quemados, es decir, huesos calentados hasta una temperatura demasiado elevada como para que provinieran de un incendio natural. Así que tal vez se tratara de restos de animales cocinados en una hoguera o un hogar.
H. erectus fue una especie de enorme éxito, no sólo por el tamaño de su población, sino también por su longevidad. Persistió durante 1,5 millones de años, desapareciendo del registro fósil hace unos 300.000 años. Sin embargo, es posible que dejara dos descendientes: H. heidelbergensis y H. neanderthalensis, conocidos respectivamente como «H. sapiens arcaico» y el famoso «hombre de Neanderthal». Ambos son clasificados a veces como subespecies (poblaciones diferenciadas pero que podían cruzarse) de H. sapiens, aunque no sabemos con certeza si contribuyeron de alguna manera al acervo genético de los humanos modernos.
H. heidelbergensis apareció hace medio millón de años, y vivió en lo que hoy es Alemania, Grecia y Francia, además de África. Presenta una mezcla de caracteres humanos modernos y de H. erectus. Los neandertales aparecieron algo más tarde, hace unos 230.000 años, y vivieron en toda Europa y en Oriente Medio. Tenían el cerebro grande (más incluso que los humanos modernos), y eran unos excelentes fabricantes de utensilios y buenos cazadores. Algunos esqueletos llevan trazas del pigmento ocre y están acompañados de «ofrendas de enterramiento», por ejemplo huesos de animales y utensilios. Esto sugiere que los neandertales enterraban a sus muertos con ceremonia, lo que quizá sea la primera indicación de una religión humana.
Pero hace unos 28.000 años, los fósiles de neandertal desaparecen. Cuando yo estudiaba, me enseñaron que simplemente habían evolucionado hacia los humanos modernos. Esta idea hoy nos parece errónea. Lo que realmente ocurriera con esta especie es tal vez la mayor incógnita de la evolución humana. Su desaparición podría estar asociada a la expansión de otra forma originaria de África: Homo sapiens. Como hemos visto, hace más o menos 1,5 millones de años H. erectus se había dispersado desde África hasta llegar incluso a Indonesia. Y dentro de esta especie había diferentes «razas», es decir, poblaciones que diferían en alguno de sus caracteres. (H. erectus de China, por ejemplo, tenía incisivos en forma de pala que no aparecen en otras poblaciones.) Entonces, hace unos 60.000 años, todas las poblaciones de H. erectus desaparecieron de repente y fueron reemplazadas por fósiles «anatómicamente modernos» de H. sapiens, que tenían esqueletos casi idénticos a los de los humanos actuales. Los neandertales aguantaron algo más, pero por fin, tras encontrar un último reducto en cuevas con vistas al estrecho de Gibraltar, también ellos dejaron paso al moderno H. sapiens. En otras palabras, Homo sapiens aparentemente echó a un lado a todos los otros homíninos de la tierra.
¿Qué es lo que ocurrió? Hay dos teorías. La primera, denominada teoría «multirregional», propone un reemplazo evolutivo: H. erectus (y quizá también H. neanderthalensis) simplemente evolucionó hacia H. sapiens de manera independiente en varias regiones, quizá porque la selección natural actuase del mismo modo en Asia, Europa y África.
La segunda idea, la teoría de la «migración desde África»,[50] propone que el moderno H. sapiens se originó en África y luego se dispersó, reemplazando físicamente a H. erectus y a los neandertales, tal vez al competir con ellos por el alimento o simplemente matándolos.
Las pruebas genéticas y fósiles apoyan la teoría de la migración desde África. ¿Por qué? Probablemente porque todo se reduce al significado de las razas. Cuanto más tiempo hayan estado separadas las poblaciones humanas, más diferencias genéticas habrán acumulado. La hipótesis multirregional, con su división en poblaciones hace más de un millón de años, predice quince veces más diferencias genéticas entre razas que si nuestros antepasados humanos salieron de África hace sólo 60.000 años. Pero tendremos ocasión más adelante de ocuparnos de las razas.
Una población de antiguos homíninos podría haber sobrevivido a la extinción en todo el mundo de H. erectus, y quizá sea la rama más extraña del árbol de familia de los humanos. Descubiertos en 2003 en la isla de Flores en Indonesia, los individuos de H. floresiensis no tardaron en recibir el apelativo de «hobbits» debido a su menguada estatura de adultos, de apenas un metro, y su peso de tan sólo unos veinticinco kilos. Su cerebro era proporcionalmente más pequeño, del tamaño del de un australopitecino, pero sus dientes y esqueleto eran indiscutiblemente de Homo. Usaban instrumentos líticos y quizá cazaban los dragones de Komodo y los elefantes enanos que poblaban la isla. Lo asombroso del caso es que floresiensis data de hace tan sólo 18.000 años, bastante después de la desaparición de los neandertales y veinticinco siglos después de que el moderno H. sapiens llegara a Australia. La hipótesis más sólida es que floresiensis representa una población aislada de H. erectus que colonizó Flores y de algún modo quedó al margen de la expansión de H. sapiens. Aunque floresiensis fue con toda probabilidad un callejón sin salida evolutivo, es difícil no dejarse cautivar por la idea de una población reciente de humanos minúsculos que cazaban elefantes enanos con diminutas lanzas, así que los hobbits han recibido un enorme interés popular.
Pero la naturaleza de los fósiles de floresiensis se discute. Algunos sostienen que la pequeña talla del único cráneo bien preservado podría corresponder simplemente a un individuo enfermo del moderno Homo sapiens, quizá uno que sufriera de cretinismo (hipotiroidismo), un trastorno que produce cráneos y cerebros anormalmente pequeños. Aunque análisis recientes de los huesos de la muñeca apoyan la idea de que H. floresiensis fue una genuina especie de homínino, aún quedan preguntas por responder.
Si miramos ahora la colección completa de huesos, ¿qué tenemos? Sin ninguna duda, una prueba indiscutible de la evolución de los humanos a partir de antepasados con aspecto de simios. Es cierto que todavía no podemos seguir un linaje continuo desde un ancestral homínino con aspecto de simio hasta el moderno Homo sapiens. Los fósiles están dispersos en el tiempo y el espacio formando una serie de puntos que todavía tenemos que acabar de conectar genealógicamente. Cabe la posibilidad de que nunca podamos conectar todos esos puntos. Pero si se colocan en orden cronológico, como en la Figura 24, lo que aparece es justamente lo que Darwin había predicho: unos fósiles que comienzan teniendo aspecto de simios pero que con el paso del tiempo se parecen cada vez más a los humanos modernos. Es un hecho que nuestra divergencia respecto a nuestro antepasado común con los chimpancés se produjo en algún lugar de África central u oriental hace unos 7 millones de años, y que la locomoción bípeda evolucionó mucho antes que el cerebro de gran tamaño. Sabemos que durante buena parte de la evolución de los homíninos coexistieron varias especies en el tiempo, a veces incluso en la misma región. Dado el pequeño tamaño de la población humana y la improbabilidad de su fosilización (recuérdese que este proceso en general requiere que un cuerpo acabe en el agua y quede cubierto rápidamente por sedimentos), es asombroso que tengamos un registro tan bueno como el que tenemos. Parece imposible examinar los fósiles que tenemos, o mirar la Figura 25, y negar que los humanos evolucionaron.
Pero algunos todavía lo hacen. Cuando se ocupan del registro fósil humano, los creacionistas realizan las más extremadas, incluso ridículas, contorsiones mentales para evitar admitir lo evidente. En realidad prefieren evitar la cuestión. Pero cuando se los fuerza a enfrentarse a ella, se limitan a ordenar los fósiles de homíninos en lo que ven como grupos distintos, los humanos y los simios, y a afirmar que estos grupos están separados por una brecha grande e insalvable. Esto refleja su visión fundamentada en la religión según la cual, aunque algunas especies podrían haber evolucionado a partir de otras, no es éste el caso de los humanos, que fueron objeto de un acto especial de creación. Pero su disparate queda manifiesto en el hecho de que los propios creacionistas no pueden ponerse de acuerdo sobre qué fósiles son «humanos» y qué fósiles son «simios». Los especímenes de H. habilis y H. erectus, por ejemplo, los clasifican como «simios» algunos creacionistas, y como «humanos» otros. Un autor ha llegado incluso a describir un espécimen de H. erectus como simio en uno de sus libros ¡y como humano en otro![51] Nada muestra el carácter intermedio de estos fósiles mejor que la incapacidad de los creacionistas para clasificarlos con claridad.
¿Qué fue, entonces, lo que impulsó la evolución de los humanos? Siempre es más fácil documentar el cambio evolutivo que entender las fuerzas que subyacen a este cambio. Lo que vemos en el registro fósil humano es la aparición de adaptaciones complejas como la postura erguida y la modificación del cráneo, que requieren muchos cambios coordinados en la anatomía, así que no cabe duda de que el proceso implicó selección natural. Pero ¿qué tipo de selección? ¿Cuáles fueron exactamente las ventajas reproductivas de un cerebro mayor, una postura erguida y unos dientes más pequeños? Es probable que nunca lo lleguemos a saber con seguridad, que sólo podamos hacer conjeturas más o menos plausibles. Lo que sí podemos es hacer conjeturas cimentadas en la información si investigamos sobre el ambiente en el que evolucionaron los humanos. Entre hace 10 y 3 millones de años, el cambio ambiental más profundo que se produjo en África central y oriental fue la sequía. Durante este período crítico de la evolución de los homíninos, el clima se fue haciendo gradualmente más seco, y le siguieron más tarde períodos alternantes y erráticos de sequía y lluvia. (Esta información proviene del polen y polvo africanos arrastrados por el viento hasta los sedimentos oceánicos, donde quedaban preservados.) Durante los períodos secos, las selvas lluviosas cedían su puesto a un hábitat más abierto, con sabanas, sistemas herbáceos, bosque claro o incluso matorral desértico. Éste es el escenario sobre el que se representó el primer acto de la evolución del hombre.
Muchos biólogos creen que estos cambios del clima y el medio tuvieron algo que ver con el primer carácter homínino que evolucionó: la locomoción bípeda. La explicación clásica es que caminar sobre dos piernas permitía a los humanos desplazarse con mayor eficacia de una mancha de bosque a otra a través del nuevo hábitat abierto. Pero esto parece improbable, pues los estudios de la locomoción bípeda y con apoyo en los nudillos han revelado que no precisan cantidades significativamente distintas de energía. Pese a ello, hay otras razones por las que caminar erguido puede haber representado una ventaja selectiva. Por ejemplo, podría haber liberado las manos para recoger y transportar formas de alimento nuevas, como la carne y los tubérculos (esto también podría explicar los dientes más pequeños y el aumento de la destreza manual). Caminar erguido también podría haber ayudado a compensar las temperaturas más altas, pues al levantar el cuerpo del suelo se reduce el área expuesta al sol. Tenemos muchas más glándulas sudoríparas que ningún otro simio, y como el pelo interfiere con el enfriamiento producido por la evaporación del sudor, quizá esto explique nuestra condición única de «monos desnudos». Hay incluso una improbable teoría del «simio acuático» que propone que los primeros homíninos pasaban buena parte del tiempo aprovisionándose de alimentos en el agua, y que la postura erguida había evolucionado para mantener la cabeza por encima de la superficie. El libro de Jonathan Kingdon sobre el bipedismo, Lowly Origin, describe aún otras teorías. Por supuesto, estas fuerzas evolutivas no son todas mutuamente excluyentes: es posible que varias de ellas actuaran al mismo tiempo. Por desgracia, no podemos discriminar unas de otras.
Lo mismo vale para la evolución del volumen cerebral. La teoría adaptativa clásica dice que una vez que nuestras manos quedaron libres por la evolución del bipedismo, los homíninos pudieron utilizarlas para fabricar utensilios, lo que llevaría a la selección de un tamaño cerebral mayor que nos habría permitido concebir y fabricar útiles más complejos. Esta teoría goza de la ventaja de que los primeros instrumentos aparecieron más o menos cuando los cerebros comenzaron a hacerse más grandes. Pero pasa por alto otras presiones selectivas que juegan a favor de un cerebro mayor y más complejo, entre ellas el desarrollo del lenguaje, la negociación de las complejidades psicológicas de la sociedad primitiva, la planificación del futuro, y otras.
Estos misterios sobre cómo evolucionamos no deberían distraernos del hecho indisputable de que en efecto evolucionamos. Incluso sin fósiles, tenemos pruebas de la evolución humana a partir de la anatomía comparada, la embriología, nuestros caracteres vestigiales e incluso la biogeografía. Ya hemos repasado nuestros embriones con aspecto de pez, nuestros genes muertos, nuestra transitoria cubierta de pelo fetal y nuestro deficiente diseño, todo lo cual es testimonio de nuestros orígenes. El registro fósil es realmente la guinda del pastel.
Nuestra herencia genética
Si todavía no entendemos por qué la selección nos ha hecho tan distintos de otros simios, ¿podemos al menos averiguar cuántos genes y de qué tipo nos diferencian? Los genes de la «humanidad» se han convertido en algo así como el santo grial de la biología evolutiva, y son muchos los laboratorios que se dedican a buscarlos. El primer intento de encontrarlos lo realizaron en 1975 Allan Wilson y Mary-Claire King, de la Universidad de California. Sus resultados fueron sorprendentes. Tras examinar secuencias de proteínas tomadas de humanos y de chimpancés, descubrieron que por término medio sólo se diferenciaban en un 1 por 100. (Estudios más recientes no han cambiado demasiado esta cifra: la diferencia ha subido hasta 1,5 por 100.) King y Wilson llegaron a la conclusión de que existía una notable similitud genética entre nosotros y nuestros parientes más próximos, y especularon que quizá unos cambios en tan sólo unos pocos genes habían producido las grandes diferencias evolutivas entre los humanos y los chimpancés. Este resultado recibió una enorme publicidad tanto en la prensa científica como en la popular, pues parecía implicar que la «humanidad» descansara en apenas un puñado de mutaciones.
Pero estudios más recientes han puesto de manifiesto que el parecido genético con nuestros primos evolutivos no es tan grande como pensábamos. Consideremos lo siguiente. Un 1,5 por 100 de diferencia en una secuencia de proteína significa que cuando alineamos la misma proteína (por ejemplo, la hemoglobina) de los humanos y la de los chimpancés, por término medio encontramos una diferencia en sólo uno de cada cien aminoácidos. Pero las proteínas suelen estar compuestas por varios centenares de aminoácidos. Así que una diferencia de 1,5 por 100 en una proteína con trescientos aminoácidos se traduce en unas cuatro diferencias en la secuencia total de la proteína. (Por usar una analogía, si cambiamos sólo un 1 por 100 de las letras de esta página, se altera mucho más del 1 por 100 de las frases.) Esa diferencia del 1,5 por 100 entre nosotros y los chimpancés que se cita tan a menudo es realmente mayor de lo que parece: mucho más del 1,5 por 100 de nuestras proteínas diferirán en al menos un aminoácido de la secuencia de los chimpancés. Y como las proteínas son esenciales para construir y mantener nuestros cuerpos, una sola diferencia puede tener efectos sustanciales.
Ahora que por fin hemos secuenciado los genomas tanto de los chimpancés como de los humanos, hemos podido ver directamente que más del 80 por 100 de todas las proteínas compartidas por las dos especies difieren en al menos un aminoácido. Puesto que nuestros genomas contienen unos 25.000 genes que codifican proteínas, esto se traduce en una diferencia en la secuencia de unas 20.000 de ellas. No es una divergencia trivial. Obviamente, lo que nos distingue es algo más que unos pocos genes. Además, los evolucionistas moleculares han descubierto recientemente que los humanos y los chimpancés difieren no sólo en la secuencia de los genes, sino también en la presencia de genes. Más de un 6 por 100 de los genes que se encuentran en los humanos no se encuentran en ninguna forma en los chimpancés. Hay más de mil cuatrocientos genes nuevos que se expresan en los humanos pero no en los chimpancés. También diferimos de los chimpancés en el número de copias de muchos de los genes que compartimos. Por ejemplo, la amilasa, una enzima de la saliva, actúa en la boca rompiendo el almidón en azúcares digeribles. Los chimpancés sólo tienen una copia de esta enzima, mientras que los humanos tenemos entre dos y dieciséis, con una media de seis copias. Esta diferencia probablemente sea el resultado de una selección natural para ayudarnos a digerir la comida, puesto que la dieta de los ancestros humanos probablemente era mucho más rica en almidón que la de los simios frugívoros.
Si ponemos todo esto sobre la mesa, vemos que la divergencia genética entre nosotros y los chimpancés se manifiesta de varias maneras: cambios no sólo en las proteínas que producen los genes, sino también en la presencia o ausencia de genes, en el número de copias de genes, y en dónde y cuándo se expresan los genes durante el desarrollo. Ya no podemos afirmar que la «humanidad» resida únicamente en un tipo de mutaciones, o en cambios en sólo unos pocos genes clave. Pero esto no es realmente sorprendente si se piensa en los muchos rasgos que nos distinguen de nuestros parientes más cercanos. Hay diferencias no sólo en anatomía, sino también en psicología (somos los simios más amorosos, y el único cuya hembra oculta la ovulación),[52] comportamiento (los humanos forman vínculos de pareja, los otros simios no), lenguaje, y en tamaño y configuración del cerebro (hay sin duda muchas diferencias en cómo se conectan las neuronas en el cerebro). Pese a nuestro parecido general con nuestros primos primates, la evolución de los humanos a partir de un antecesor con aspecto de simio probablemente requirió cambios genéticos sustanciales.
¿Podemos decir algo sobre los genes específicos que nos han hecho humanos? Por el momento, no mucho. Con la ayuda de «escaneos» genómicos que comparan la secuencia entera de ADN de los chimpancés y los humanos, podemos escoger clases de genes que han evolucionado con rapidez en la rama humana de la divergencia. Estas clases incluyen genes involucrados en el sistema inmunitario, la formación de gametos, la muerte celular y, lo más interesante de todo, la percepción sensorial y la formación de nervios. Pero afinar hasta el nivel de genes concretos y demostrar que las mutaciones en ese gen realmente produjeron diferencias entre humanos y chimpancés ya es harina de otro costal. Hay algunos candidatos a genes de este tipo, entre ellos uno (FOXP2) que podría haber estado involucrado en la aparición del lenguaje,[53] aunque no tenemos pruebas concluyentes. Tal vez no las tengamos nunca. Las pruebas concluyentes de que un gen concreto causa diferencias entre humanos y chimpancés requeriría el traslado del gen de una especie a otra para ver qué diferencia produce, y ése no es un tipo de experimento que alguien querría intentar.[54]
La espinosa cuestión de la raza
Cuando se viaja por el mundo, enseguida se ve que los humanos de distintos lugares tienen aspecto distinto. Nadie, por ejemplo, confundiría un japonés y un finés. La existencia de tipos humanos visiblemente distintos es obvia, pero no hay en la biología mayor campo de minas que la cuestión de la raza. La mayoría de los biólogos se mantienen tan alejados de ella como pueden. Una mirada a la historia de la ciencia nos dice por qué. Desde el principio de la biología moderna, la clasificación de las razas ha ido de la mano de los prejuicios raciales. En su clasificación de las razas, Linnaeus observó en el siglo XVIII que los europeos están «gobernados por leyes», los asiáticos «gobernados por opiniones» y los africanos «gobernados por caprichos». En su soberbio libro La falsa medida del hombre, Stephen Jay Gould documenta la perversa conexión entre la biología y la raza durante el último siglo.
En respuesta a estos desagradables episodios de racismo, algunos científicos han reaccionado de manera exagerada, argumentando que las razas humanas no tienen realidad biológica, que son meros «constructos» sociopolíticos que, como tales, no son dignos de estudio científico. Pero para los biólogos, el término «raza» siempre ha sido totalmente respetable, ¡siempre y cuando no se aplique a los humanos! Las razas (también llamadas «subespecies» o «ecotipos») son simplemente poblaciones de una especie que están geográficamente separadas y que difieren genéticamente en uno o más caracteres. Hay muchas razas de plantas y animales, incluidas las poblaciones de ratón de campo que difieren sólo en el color del pelaje, las poblaciones de gorriones que difieren en el tamaño y el canto, y las razas de plantas que difieren en la forma de las hojas. Siguiendo esta definición, Homo sapiens claramente tiene razas. Y el hecho de que las tengamos es otra indicación de que no somos distintos de las otras especies que han evolucionado.
La existencia de razas distintas en los humanos muestra que nuestras poblaciones estuvieron separadas el tiempo suficiente para que se produjera alguna divergencia genética. Pero ¿cuánta divergencia? Y ¿se ajusta a lo que indican los fósiles sobre nuestra expansión desde África? ¿Qué tipo de selección impulsó esas diferencias?
Como podíamos esperar de la evolución, la variación física de los humanos se produce en grupos anidados unos en otros, y pese a los esforzados intentos de algunos de crear divisiones formales de razas, es completamente arbitrario dónde exactamente se sitúa la línea que demarca una raza particular. No hay fronteras marcadas: el número de razas reconocidas por los antropólogos ha ido variando entre tres y más de treinta. Una mirada a los genes muestra con mayor claridad si cabe la falta de fronteras marcadas entre las razas: prácticamente toda la variación genética revelada por las modernas técnicas moleculares muestra sólo una correlación débil con las combinaciones clásicas de caracteres físicos que se suelen utilizar para determinar la raza, como el color de la piel y el tipo de cabello.
Las pruebas genéticas directas, acumuladas durante las tres últimas décadas, muestran que sólo alrededor del 10 al 15 por 100 de toda la variación genética de los humanos está representada por diferencias entre «razas» que puedan reconocerse por diferencias en la apariencia física. El resto de la variación genética, del 85 al 90 por 100, se produce entre individuos dentro de las razas.
Lo que esto significa es que las razas no muestran diferencias de todo o nada en las formas de los genes (alelos) que poseen, sino que por lo general tienen los mismos alelos, pero con frecuencias distintas. El gen del grupo sanguíneo ABO, por ejemplo, tiene tres alelos: A, B y O. Casi todas las poblaciones humanas tienen estas tres formas, pero se encuentran con frecuencias distintas en distintos grupos. El alelo O, por ejemplo, tiene una frecuencia de 54 por 100 en los japoneses, 64 por 100 en los fineses, 74 por 100 en los !kung sudafricanos y 85 por 100 en los navajos. Éste es un caso típico de la clase de diferencias que vemos en el ADN: no se puede diagnosticar el origen de una persona a partir de un solo gen, sino que hay que examinar una combinación de muchos genes.
En el nivel genético, por tanto, los seres humanos son notablemente parecidos. Eso es precisamente lo que cabe esperar si los humanos modernos salieron de África hace sólo de 60.000 a 100.000 años. No ha habido mucho tiempo para la divergencia genética, aunque nos hayamos diseminado por las cuatro esquinas del mundo, disgregándonos en varias poblaciones muy alejadas que se han mantenido genéticamente aisladas hasta décadas recientes.
¿Significa esto que podemos ignorar las razas humanas? No. Estas conclusiones no significan que las razas sean meros constructos mentales o que las pequeñas diferencias genéticas entre ellas carezcan de interés. Algunas diferencias raciales nos ofrecen indicios claros de las presiones evolutivas que actuaron en distintas regiones, y pueden resultar útiles en la medicina. La anemia falciforme, por ejemplo, es más común en los hombres de raza negra cuyas antepasados provengan del África ecuatorial. Como los portadores de la mutación de esta anemia poseen cierta resistencia a la malaria provocada por Plasmodium falciparium (la forma más mortífera de esta enfermedad), es probable que la elevada frecuencia de esta mutación en las poblaciones africanas y derivadas de África sea consecuencia de la selección natural en respuesta a la malaria. La enfermedad de Tay-Sachs es un trastorno genético mortal común entre los judíos asquenazíes y los cajunes de Luisiana, que probablemente ha alcanzado estas altas frecuencias por deriva genética en poblaciones ancestrales pequeñas. Conocer la etnia de un paciente es de enorme utilidad para el diagnóstico de estas y otras enfermedades congénitas. Además, las diferencias en las frecuencias alélicas entre los grupos raciales significa que la búsqueda de donantes de órganos apropiados, que requieren la coincidencia entre varios «genes de compatibilidad», debería tener en cuenta la raza.
La mayoría de las diferencias genéticas entre razas son triviales. Otras, sin embargo, como las diferencias físicas entre un japonés y un finés, un masai y un inuit, son muy manifiestas. Nos encontramos, pues, ante la interesante situación de que las diferencias globales en las secuencias de genes entre los pueblos son pequeñas, pero esos mismos grupos presentan diferencias drásticas en diversos rasgos visualmente manifiestos, como el color de la piel, el color del cabello, la forma del cuerpo y la forma de la nariz. Estas obvias diferencias físicas no son características del genoma en su conjunto. ¿Por qué la pequeña cantidad de divergencia que se ha producido entre las poblaciones humanas se ha centrado en una serie de rasgos visualmente llamativos?
Algunas de estas diferencias tienen sentido como adaptaciones a los distintos ambientes en los que se han encontrado los humanos. El color de piel más oscuro de los grupos tropicales probablemente ofrezca una mayor protección frente a la intensa luz ultravioleta que produce melanomas letales, mientras que la piel pálida de los grupos de latitudes más altas facilita la penetración de la luz necesaria para la síntesis de la esencial vitamina D, que ayuda a prevenir el raquitismo y la tuberculosis.[55] Pero ¿qué sentido tiene la forma de los ojos de los asiáticos, o la nariz más larga de los caucásicos? Estos rasgos no tienen ninguna relación con el medio. Para algunos biólogos, la existencia de una mayor variación entre razas en los genes que afectan a la apariencia física, que es algo que pueden evaluar fácilmente las parejas potenciales, apunta a un sola causa: la selección sexual.
Aparte de la pauta característica de variación genética, hay otras razones para considerar la selección sexual como una poderosa fuerza impulsora de la evolución de las razas. Somos únicos entre las especies por haber desarrollado culturas complejas. El lenguaje nos ha proporcionado la notoria capacidad de divulgar ideas y opiniones. Un grupo de humanos puede cambiar su cultura mucho más rápido de lo que puede evolucionar genéticamente. Pero el cambio cultural también puede producir cambios genéticos. Imaginemos que la expansión de una idea o moda implica preferencias sobre la apariencia de la pareja. Una emperatriz de Asia, por ejemplo, podría tener debilidad por los hombres de pelo negro y lacio y ojos almendrados. Al crear una moda, su preferencia se extiende culturalmente a todas sus súbditas, y, mira por dónde, con el tiempo los individuos de pelo rizado y ojos redondos son reemplazados por los de pelo lacio y ojos almendrados. Es esta «coevolución gen-cultura», la idea de que un cambio en el entorno cultural conduce a nuevos tipos de selección de los genes, lo que hace especialmente atractiva la idea de la selección sexual de las diferencias físicas.
Además, la selección sexual a menudo puede actuar con increíble rapidez, lo que la convierte en un candidato ideal para impulsar la rápida diferenciación evolutiva de los rasgos físicos que se produjeron desde la más reciente migración de nuestros antepasados desde África. Por supuesto, todo esto es sólo especulación, y casi imposible de contrastar, pero al menos potencialmente explica ciertas diferencias desconcertantes entre grupos.
No obstante, la mayor parte de la controversia sobre la raza no se centra en las diferencias físicas entre poblaciones, sino en diferencias de conducta. ¿Ha hecho la evolución que ciertas razas sean más inteligentes, más atléticas o más astutas que otras? Llegados aquí se impone la cautela, pues en este ámbito las afirmaciones sin un fundamento firme pueden acabar brindándole crédito científico al racismo. Pero ¿qué dicen los datos científicos? No dicen casi nada. Aunque las distintas poblaciones puedan tener conductas distintas, coeficientes intelectuales distintos o distintas capacidades, es difícil descartar la posibilidad de que estas diferencias sean un producto no genético de diferencias ambientales o culturales. Si deseamos determinar si ciertas diferencias entre razas tienen una base en los genes, tenemos que descartar estas influencias. Los estudios de este tipo requieren experimentos controlados: quitar de sus padres a unos bebés de distintas etnias y criarlos a todos en entornos idénticos (o aleatorios). Las diferencias conductuales que quedasen serían genéticas. Como estos experimentos no son éticos, no se han realizado de manera sistemática, pero las adopciones entre culturas distintas muestran de forma anecdótica que las influencias culturales sobre el comportamiento son fuertes. Como bien observa el psicólogo Steven Pinker: «Un niño adoptado de una parte tecnológicamente subdesarrollada del mundo no tiene ningún problema en adaptarse a la sociedad moderna». Esto sugiere, al menos, que las razas no presentan grandes diferencias innatas en el comportamiento.
Mi impresión, que no pasa de una especulación bien cimentada en la información, es que las razas humanas son demasiado jóvenes para haber producido por evolución diferencias importantes en inteligencia o conducta. Y que tampoco hay buenas razones para pensar que la selección natural o sexual haya favorecido este tipo de diferencias. En el capítulo siguiente examinaremos muchos comportamientos «universales» que se ven en todas las sociedades humanas, comportamientos como el lenguaje simbólico, el miedo a los extraños durante la infancia, la envidia, el cotilleo y la costumbre de hacer regalos. Si estos universales tienen alguna base genética, su presencia en cada sociedad da un mayor peso a la idea de que la evolución no ha producido una divergencia psicológica sustancial entre los grupos humanos.
Aunque ciertos rasgos como el color de la piel y el tipo de pelo han divergido entre poblaciones, al parecer se trata de casos especiales, impulsados por diferencias ambientales entre localidades o por selección sexual de la apariencia externa. Los datos de ADN muestran que, en términos generales, las diferencias genéticas entre poblaciones humanas son pequeñas. Decir que todos somos iguales bajo la piel es algo más de un tópico reconfortante. Y eso es precisamente lo que cabe esperar si se tiene en cuenta el breve período de evolución desde nuestro más reciente origen en África.
Pero ¿y ahora?
La selección no parece haber producido grandes diferencias entre razas, pero sí algunas intrigantes diferencias entre poblaciones dentro de grupos étnicos. Como estas poblaciones son bastante jóvenes, lo que tenemos es una prueba clara de que la selección ha actuado en los humanos en tiempos recientes.
Uno de los casos tiene que ver con nuestra capacidad para digerir la lactosa, un azúcar que se encuentra en la leche. Una enzima llamada lactasa descompone este azúcar en glucosa y galactosa, que son absorbidos más fácilmente. Como es natural, nacemos con la capacidad de digerir la leche, pues ése ha sido desde siempre el principal alimento de los bebés. Pero una vez destetados, de manera gradual dejamos de producir lactasa. Al final, muchos de nosotros perdemos completamente la capacidad de digerir la lactosa, es decir, adquirimos una «intolerancia a la lactosa» que nos hace propensos a sufrir diarreas, hinchazón y retortijones después de comer un producto lácteo. La desaparición de la lactasa después del destete probablemente sea el resultado de la selección natural: nuestros antepasados más antiguos no tenían ninguna fuente de leche una vez acabado el amamantamiento, así que ¿para qué producir una costosa enzima que ya no se necesita?
Pero en algunas poblaciones humanas, los individuos siguen produciendo lactasa durante toda la edad adulta, lo que les proporciona una buena fuente de nutrición que no está disponible para otros. Lo interesante del caso es que la persistencia de la lactasa se encuentra sobre todo en las poblaciones que fueron, o todavía son, pastorales, o sea que crían vacas. Se incluyen aquí varias poblaciones europeas y de Oriente Medio, y algunas africanas como los masai y los tutsi. Los análisis genéticos muestran que la persistencia de la lactasa en estas poblaciones depende de un simple cambio en el ADN que regula la enzima, que se mantiene activa después de la infancia. Este gen tiene dos alelos, la forma «tolerante» (activo) y la «intolerante» (inactivo), que difieren en una sola letra del código de ADN. La frecuencia del alelo tolerante se correlaciona bien con el uso de vacas por las poblaciones: es alta (50 a 90 por 100) en las sociedades pastorales de Europa, Oriente Medio y África, y muy baja (1 a 20 por 100) en las poblaciones asiáticas y africanas que dependen de la agricultura en lugar de la leche.
Las observaciones arqueológicas muestran que los humanos comenzaron a domesticar vacas hace entre 7.000 y 9.000 años en Sudán, y la práctica se extendió por el África subsahariana y Europa unos pocos miles de años más tarde. La parte más gratificante de esta historia es que podemos determinar, a partir de la secuenciación de ADN, cuándo apareció por mutación el alelo «tolerante». Ese tiempo, hace de 3.000 a 8.000 años, encaja notablemente bien con el aumento del pastoralismo. Y lo que es mejor, el ADN extraído de esqueletos de europeos de hace 7.000 años muestra que eran intolerantes a la lactosa, como es de esperar si se tiene en cuenta que todavía no eran una sociedad pastoral.
La evolución de la tolerancia a la lactosa es otro ejemplo espléndido de coevolución gen-cultura. Un cambio puramente cultural (la cría de vacas, quizá para obtener carne) produjo una nueva oportunidad para la evolución: la capacidad de utilizar esas vacas para obtener leche. Dada esta súbita disponibilidad de un nuevo y nutritivo alimento, los antepasados que poseían el gen de la tolerancia debían tener una ventaja reproductiva considerable respecto a los que tenían el gen intolerante. De hecho, podemos calcular esta ventaja observando con qué velocidad aumentó la frecuencia del gen de la tolerancia en las poblaciones modernas. El resultado es que los individuos tolerantes deben haber producido, por término medio, de 4 a 10 por 100 más descendientes que quienes eran intolerantes. Ésta es una selección bastante fuerte.[56]
Cualquiera que dé clases de evolución humana recibe inevitablemente la siguiente pregunta: ¿todavía estamos evolucionando? Los ejemplos de la tolerancia a la lactosa y la duplicación del gen de la amilasa muestran que la selección sin duda ha actuado durante los últimos miles de años. Pero ¿todavía actúa en la actualidad? Es difícil dar una respuesta clara. Es obvio que muchos tipos de selección que supusieron desafíos para nuestros antepasados ya no se aplican: las mejoras en nutrición, higiene y sanidad han acabado con muchas enfermedades y trastornos que mataban a nuestros antepasados y han eliminado fuentes de selección natural en otro tiempo potentes. Como dice el genetista británico Steve Jones, hace quinientos años un niño británico tenía sólo un 50 por 100 de probabilidad de sobrevivir hasta la edad reproductora, una cifra que en la actualidad ha ascendido hasta el 99 por 100. Y entre quienes han sobrevivido, la intervención médica ha permitido que lleven vidas normales a muchas personas que la selección natural habría eliminado sin piedad durante la mayor parte de nuestra historia evolutiva. ¿Cuánta gente con mala vista o mala dentadura, incapacitados para cazar o masticar, hubieran muerto en la sabana africana? (Yo seguro que hubiera estado entre los incapacitados.) ¿Cuántos de nosotros hemos tenido infecciones que, sin antibióticos, nos hubieran matado? Es probable que, a causa del cambio cultural, nos estemos deteriorando genéticamente de varias maneras. Es decir, genes que en otro tiempo eran perjudiciales han dejado de ser tan malos (podemos compensar unos «malos» genes con un par de gafas o un buen dentista), de manera que persisten en la población.
Y al contrario, genes que en otro tiempo fueron buenos pueden tener ahora efectos destructivos debido a cambios culturales. Nuestra pasión por los dulces y las grasas, por ejemplo, seguramente fue adaptativa para nuestros antepasados, para quienes estas delicias eran una fuente de energía valiosa pero poco frecuentes.[57] Pero ahora que estos alimentos son abundantes, nuestra herencia genética nos induce problemas de deterioro de los dientes, de obesidad y cardiovasculares. Nuestra tendencia a acumular la grasa de los alimentos ricos en energía también debió ser adaptativa cuando la variación en la abundancia local de alimentos producía situaciones de exceso o hambruna, en las que los individuos que podían almacenar calorías para los tiempos de vacas flacas tenían una ventaja selectiva.
¿Significa esto que estamos desevolucionando? Hasta cierto punto, sí, pero probablemente también nos estemos adaptando mejor a nuestro entorno moderno, que crea nuevos tipos de selección. Debemos recordar que mientras haya personas que mueran antes de acabar de reproducirse, y mientras algunas personas dejen más descendencia que otras, hay espacio para que la selección natural nos cambie. Y si existe variación genética que afecte a nuestra capacidad para sobrevivir y dejar hijos, promoverá el cambio evolutivo. Esto sin duda está ocurriendo en la actualidad. Aunque la mortalidad prerreproductora sea baja en algunas poblaciones occidentales, sigue siendo alta en otros lugares, sobre todo en África, donde la mortalidad infantil puede superar el 25 por 100. Y esa mortalidad a menudo tiene su causa en enfermedades infecciosas como el cólera, la fiebre tifoidea y la tuberculosis. Otras enfermedades, como la malaria y el sida, siguen matando a muchos niños y adultos en edad reproductora.
Las fuentes de mortalidad están ahí, como también los genes para aliviarlas. Los alelos variantes de ciertas enzimas, por ejemplo de la hemoglobina (en especial el alelo falciforme), confieren resistencia a la malaria. Y hay un gen mutante, un alelo llamado CCR5-Δ32, que proporciona a sus portadores una fuerte protección frente a la infección por el virus del sida. Podemos predecir que si el sida continúa siendo una fuente significativa de mortalidad, la frecuencia de este alelo aumentará en las poblaciones afectadas. Eso es la evolución, igual que lo es la resistencia a los antibióticos en las bacterias. Y sin duda existen otras fuentes de mortalidad que todavía no entendemos adecuadamente: toxinas, contaminación, estrés y otras por el estilo. Si algo hemos aprendido de los experimentos de cruzamientos es que prácticamente todas las especies tienen variación genética para responder a casi cualquier forma de selección. Lenta pero inexorablemente, nuestro genoma se adapta a muchas nuevas fuentes de mortalidad. Pero no a todas. Las afecciones que tienen causas tanto genéticas como ambientales, como la obesidad, la diabetes y las enfermedades cardiovasculares, pueden no responder a la selección porque la mortalidad que producen se da sobre todo después de que las víctimas hayan dejado de reproducirse. La supervivencia de los más dotados es acompañada por la supervivencia de los más gordos.[58]
Pero no es la resistencia a las enfermedades lo que interesa a la gente, por importante que sea. Lo que quieren saber es si los humanos se están haciendo más fuertes, más listos o más guapos. Eso, naturalmente, depende de si esos rasgos se asocian a una reproducción diferencial, y eso lo desconocemos. Tampoco es que importe demasiado. En nuestra cultura, que cambia con tanta rapidez, los progresos sociales mejoran nuestras capacidades mucho más que cualquier cambio en nuestros genes, a no ser, claro está, que decidamos intervenir en nuestra evolución por medio de la manipulación genética, por ejemplo preseleccionando espermatozoides y óvulos favorables.
Así pues, la lección que nos enseña el registro fósil, combinada con los descubrimientos más recientes de genética humana, confirman que somos mamíferos evolucionados, mamíferos orgullosos y de éxito, sin duda, pero mamíferos al fin y al cabo, construidos por los mismos procesos que transformaron cualquier otra forma de vida durante los últimos miles de millones de años. Como todas las especies, no somos un producto final de la evolución, sino una obra en progreso, por mucho que nuestro progreso genético sea lento. Y aunque hemos llegado muy lejos desde los simios ancestrales, las marcas de nuestra herencia todavía se manifiestan. Gilbert y Sullivan bromeaban diciendo que no éramos más que unos monos depilados; Darwin no era tan gracioso, pero sí mucho más lírico y veraz:
He ofrecido todas las pruebas que mi capacidad me ha permitido; y debemos reconocer, según me parece a mí, que el hombre con todas sus nobles cualidades, con su compasión hacia los más degradados, con su benevolencia no sólo hacia los otros hombres sino hacia la más humilde criatura; con su intelecto, que parece divino y ha penetrado en los movimientos y la constitución del sistema solar; con todos estos elevados poderes, todo hombre lleva todavía en su estructura corporal el sello indeleble de su humilde origen.