
Capítulo 6
No podemos suponer, por ejemplo, que los machos de las aves del paraíso o los pavos reales realicen tantos esfuerzos para levantar, extender y hacer vibrar sus hermosas plumas ante las hembras sin ningún propósito.
CHARLES DARWIN
Pocos animales hay en la naturaleza más deslumbrantes que el macho del pavo real en plena exhibición, con su iridiscente cola verde y azul tachonada de ocelos, abierta como un abanico en pleno esplendor tras su cuerpo azul brillante. Pero esta ave parece violar todos los aspectos del darwinismo, pues los mismos caracteres que la hacen hermosa constituyen malas adaptaciones para la supervivencia. Esa larga cola provoca problemas aerodinámicos para volar, como bien sabrá cualquiera que haya visto un pavo real intentado alzar el vuelo. No hay duda de que esto debe dificultarle el ascenso a su percha en los árboles donde pasa la noche, o la huida de sus depredadores, sobre todo durante los monzones, cuando la cola mojada se convierte en una auténtica carga. Además, los colores brillantes atraen a los depredadores, especialmente por comparación con las hembras, que tienen la cola corta y el cuerpo de un color verde pardusco apagado que les sirve de camuflaje. Los machos tienen que desviar buena parte de su energía metabólica a esta sorprendente cola, que crece completamente de nuevo cada año.
El plumaje del pavo real no sólo parece inútil, también un impedimento. ¿Cómo puede ser una adaptación? Y si los individuos con este plumaje dejan más genes, como cabe esperar si tal atuendo evolucionó por selección natural, ¿por qué las hembras no son igual de resplandecientes? En una carta dirigida al biólogo norteamericano Asa Gray en 1860, Darwin se quejaba amargamente de esta cuestión: «Aún recuerdo los tiempos en que el solo pensamiento del ojo me producía escalofríos, pero he superado esa fase de lamentación y ahora suelen ser los detalles insignificantes de estructura los que me incomodan. La visión de una pluma de la cola de un pavo real, cada vez que la observo, ¡me marea!».
Los enigmas como el de la cola del pavo real son abundantes. Ahí está, sin ir más lejos, el extinto alce irlandés (mal nombre para un animal que ni era alce ni exclusivo de Irlanda; fue el mayor de los ciervos jamás descrito y vivió en toda Europa y Asia). Los machos de esta especie, que desaparecieron hace sólo diez mil años, ostentaban una enorme cornamenta con una envergadura de ¡más de 3,5 metros de punta a punta! Con un peso de unos cuarenta kilos, se asentaba en un insignificante cráneo de unos dos kilos y medio. Qué presión no debía causar: como andar todo el día con un adolescente sobre la cabeza. Para colmo de males, como pasaba con la cola del pavo real, la cornamenta se rehacía completamente cada año.
Aparte de los caracteres llamativos, hay conductas extrañas que se observan en uno solo de los sexos. Los machos de las ranas túngara de América Central usan sus sacos vocales inflables para cantar cada noche unas largas serenatas. Sus canciones atraen la atención de las hembras, pero también de los murciélagos y las moscas hematófagas (que se alimentan de sangre), que depredan sobre los machos de rana mucho más que sobre las hembras, que no cantan. En Australia, los machos de las aves de emparrado construyen con palitos unos grandes y extraños «emparrados» que, dependiendo de la especie, tienen la forma de un túnel, de una seta o de una tienda. Estas estructuras están decoradas con todo tipo de ornamentos: flores, conchas de caracoles, bayas, vainas de semillas y, cuando cerca viven humanos, tapones de botellas, trozos de vidrio y papel de aluminio. Tardan horas en construir estos emparrados, a veces días (algunos llegan a medir tres metros de alto por uno y medio de ancho), pero no los usan como nido. ¿Por qué se toman tanto trabajo los machos?
No tenemos que limitamos a especular, como hizo Darwin, que estos caracteres reducen la supervivencia. Durante los últimos años los científicos han medido el coste que pueden llegar a tener. El macho del obispo de collar rojo[34] es de color negro brillante, con un collar y una mancha en la cabeza de un intenso color carmesí, y unas larguísimas plumas en la cola, más o menos el doble de largas que su cuerpo. Cualquiera que vea el macho en pleno vuelo, luchando por avanzar mientras su larga cola se bambolea a su espalda, no puede sino preguntarse por esa cola. Sarah Pryke y Steffan Andersson, de la Universidad Göterborg, de Suecia, capturaron un grupo de machos en América del Sur y les cortaron la cola, reduciéndola en unos dos centímetros y medio en un grupo y unos diez en otro. Por medio de varias recapturas realizadas a lo largo del verano, descubrieron que los machos con la cola larga perdían una cantidad de peso significativamente mayor que los de cola corta. No cabe duda de que esas largas colas suponen una desventaja.
También lo son los colores brillantes, como se ha demostrado mediante un ingenioso experimento con el lagarto de collar. En este lagarto de unos treinta centímetros que vive en el oeste de Estados Unidos, los dos sexos tienen un aspecto muy distinto: los machos exhiben un cuerpo turquesa, con la cabeza amarilla, collar negro y manchas blancas y negras, mientras que las hembras son de color gris pardo con pocas manchas. Para contrastar la hipótesis de que el color brillante de los machos atrae a más depredadores, Jerry Husak y sus colegas de la Universidad Estatal de Oklahoma pusieron en el desierto unos modelos de arcilla pintados como los machos o como las hembras. La arcilla blanda conservaba las marcas de los mordiscos de los depredadores que confundían los modelos con animales de verdad. Tras sólo una semana, treinta y cinco de los cuarenta modelos chillones de los machos presentaban marcas de mordiscos, la mayoría de serpientes y aves, mientras que ninguno de los cuarenta modelos sosos de las hembras había sido atacado.
Los caracteres que difieren entre los machos y las hembras de una especie, como la cola, el color y las canciones, reciben el nombre de dimorfismos sexuales, del griego «dos formas». (La Figura 23 muestra varios ejemplos.) Los biólogos han encontrado repetidas veces que los caracteres sexualmente dimórficos en los machos parecen violar la teoría de la evolución, pues derrochan tiempo y energía y reducen la supervivencia. Los coloridos machos de los gupis acaban siendo comidos más a menudo que las hembras, que pasan más desapercibidas. El macho de la collalba gris, un pájaro mediterráneo, construye laboriosamente unos grandes hitos con piedras apiladas en distintos lugares, apilando en ellos hasta cincuenta veces su propio peso en guijarros a lo largo de un par de semanas. El macho del gallo de las artemisas realiza una elaborada exhibición, contoneándose arriba y abajo por la pradera, y aleteando mientras produce unos fuertes sonidos que emanan de dos grandes sacos vocales.[35] Tanto lucimiento puede quemar una enorme cantidad de energía para estas aves. Si la selección es responsable de esos caracteres, y dada su complejidad debería serlo, necesitamos explicar de qué modo.
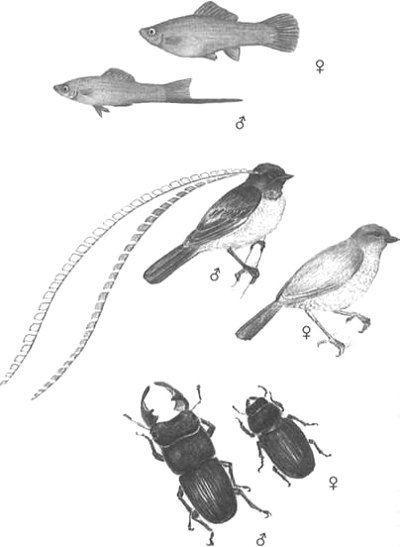
FIGURA 23. Ejemplo de dimorfismo sexual donde se muestran marcadas diferencias en el aspecto de machos y hembras. Arriba: el cola de espada (Xiphophorus helleri); en medio: ave del paraíso del rey de Sajonia (Pteridophora alberti), cuyos machos poseen elaborados ornamentos en la cabeza de color azul celeste por un lado y pardos por el otro; abajo: un ciervo volante (Aegus formosae).
Las soluciones
Antes de Darwin, el dimorfismo sexual era un misterio. Entonces, igual que ahora, los creacionistas no podían explicar por qué un diseñador sobrenatural había producido caracteres en un sexo, y sólo en uno, que perjudicaban a su supervivencia. Como gran explicador de la diversidad de la naturaleza, Darwin naturalmente estaba ansioso por entender cómo habían evolucionado aquellos rasgos en apariencia inútiles. Por fin comprendió la clave para explicarlos: cuando los caracteres difieren entre los machos y las hembras de una especie, las elaboradas conductas, estructuras y ornamentos están casi siempre restringidos a los machos.
A estas alturas, el lector ya se habrá imaginado cómo evolucionaron estos caracteres tan costosos. Hay que recordar que en realidad la moneda de cambio de la selección no es la supervivencia, sino el éxito reproductor. Tener una cola estrafalaria o un canto seductor no ayuda a sobrevivir, pero puede incrementar la probabilidad de dejar descendencia, y así es como surgieron estas conductas y rasgos ostentosos. Darwin fue el primero en reconocer este compromiso, y acuñó un nombre para el tipo de selección responsable de los caracteres sexuales dimórficos: selección sexual. La selección sexual es sencillamente la selección que aumenta las probabilidades de que un individuo consiga pareja. No es más que un caso particular de la selección natural, pero merece un capítulo aparte por la forma única en que actúa y por las soluciones aparentemente no adaptativas que produce.
Los caracteres seleccionados sexualmente evolucionan sólo cuando compensan la reducción de la supervivencia con un aumento de la reproducción. Quizá los obispos de collar rojo con una cola más larga no escapen muy bien de los depredadores, pero las hembras los prefieren como pareja. Los ciervos con mayor cornamenta se las ven y se las desean para sobrevivir con esa carga metabólica, pero quizá así ganan más peleas durante las berreas, y por tanto dejan más descendencia.
La selección sexual se presenta en dos formas. La primera, ejemplificada por las enormes astas del alce irlandés, es la competencia directa entre machos por las hembras. La otra, la que produce la larga cola del obispo de collar rojo, es la selectividad de las hembras entre los machos posibles. La competencia entre machos (lo que Darwin, con su terminología a menudo pugnaz, llamaba «ley de la batalla») es la más fácil de entender. Como Darwin hizo notar, «es una certeza que en casi todos los animales se produce una lucha entre los machos por la posesión de las hembras». Cuando los machos de una especie luchan entre sí de manera directa, ya sea haciendo chocar las cornamentas como en los ciervos, intentado clavarse los cuernos como en el caso del ciervo volante, golpeándose la cabeza como en las moscas de ojos pedunculados o mediante enfrentamientos sangrientos, como en los enormes elefantes marinos, consiguen llegar a las hembras echando a un lado a sus competidores. La selección favorecerá cualquier carácter que promueva tales victorias en tanto en cuanto el aumento de la posibilidad de obtener pareja iguale y supere la reducción de la supervivencia. Este tipo de selección produce armamentos: armas más fuertes, cuerpo de mayor tamaño o cualquier cosa que ayude a un macho a ganar en un enfrentamiento físico.
En cambio, los caracteres como los colores intensos, los ornamentos, los emparrados y las exhibiciones de cortejo están moldeados por el segundo tipo de selección sexual, la elección del macho. A los ojos de las hembras, no todos los machos son iguales. Algunos rasgos y conductas de los machos les resultan más atractivos que otros, así que los genes que producen esos caracteres se acumulan en las poblaciones. Existe aquí también un elemento de competencia entre los machos, pero es indirecto: los machos que ganan tienen las voces más altas, los colores más vivos, las hormonas más atrayentes, las exhibiciones más sensuales… pero, en contraste con la competencia entre machos, el ganador lo deciden las hembras.
En ambos tipos de selección sexual, los machos compiten por las hembras. ¿Por qué no al revés? Enseguida veremos que todo depende de la diferencia de tamaño entre dos células diminutas: el espermatozoide y el óvulo.
Pero ¿es realmente cierto que los machos que ganan las batallas, o que están más ornamentados, o que realizan las mejores exhibiciones, se llevan más parejas? Si no fuera así, la teoría entera de la selección sexual se caería por su propio peso.
La realidad es que las pruebas y observaciones de que disponemos apoyan sin resquicios la teoría. El elefante marino septentrional de la costa pacífica de América del Norte presenta un dimorfismo sexual extremo en el tamaño. Las hembras no miden más de unos tres metros de largo y pesan unos 700 kilos, mientras que los machos pueden ser el doble de grandes y llegan a pesar hasta 2.730 kilos, o sea, que son más grandes y pesan más del doble que un Volkswagen. Además, son políginos: los machos se aparean con más de un hembra durante la misma época de cría. Aproximadamente un tercio de los machos controlan un harén de hembras con las que se aparean (¡hasta un centenar de parejas para un macho alfa!), mientras que el resto de los machos están condenados a la soltería. Quién gana y quién pierde en la lotería del apareamiento es algo que se decide mediante feroces combates entre los machos antes incluso de que las hembras se arrastren hasta la playa. En estos sangrientos combates los grandes machos golpean sus enormes masas unos contra otros, se infligen profundas heridas en el cuello con sus colmillos, y el resultado es una jerarquía de dominancia en cuya cima se hallan los machos más grandes. Cuando llegan las hembras, los machos dominantes las conducen hasta sus harenes y rechazan a los machos rivales. En un año determinado, la mayoría de los cachorros son hijos de unos pocos machos más grandes.
Esto es competencia entre machos, pura y simple, y el premio es la reproducción. Es fácil ver que, con este sistema de apareamiento, la selección sexual promoverá la evolución de machos grandes y fieros, pues son los más grandes, no los pequeños, los que pasan sus genes a la siguientes generación. (Las hembras, que no tienen que luchar, supuestamente tienen un peso cercano al óptimo para la reproducción.) El dimorfismo sexual del tamaño corporal de muchas especies, incluida la nuestra, podría deberse a la competencia entre los machos por las hembras.
Los machos de las aves a menudo compiten con fiereza por el espacio. En muchas especies, los machos atraen a las hembras sólo cuando controlan un trozo de tierra con buena vegetación y adecuado para la nidificación. Una vez conseguida su parcela, los machos la defienden con exhibiciones visuales y vocales, y por supuesto con ataques directos a los machos que se acerquen demasiado. Muchos de los cantos de pájaro que nos deleitan los oídos son en realidad amenazas, advertencias dirigidas a otros machos para que se mantengan alejados.
El turpial de hombros rojos de América del Norte defiende territorios en hábitats abiertos, por lo general zonas pantanosas de agua dulce. Igual que el elefante marino, esta especie es polígina, y algunos machos tienen hasta quince hembras nidificando en su territorio. Muchos otros machos, los llamados «flotantes», se quedan sin pareja. Los flotantes intentan constantemente invadir los territorios establecidos para copular de extranjís con las hembras, así que los machos residentes están siempre muy ocupados intentando echarlos. Hasta una cuarta parte de su tiempo se la pasa el macho protegiendo activamente su territorio. Además de patrullar por su terreno, los machos de turpial defienden su territorio cantando complejas canciones y realizando exhibiciones amenazadoras con su ornamento epónimo, una charretera de color rojo vivo que llevan en el hombro. (Las hembras son de color pardo, a veces con una charretera pequeña y vestigial.) Las charreteras no están ahí para atraer a las hembras, sino para amenazar a otros machos en las batallas por el territorio. Si experimentalmente se borran las charreteras de los machos pintándolas de negro, el 70 por 100 de los machos pierden su territorio, en comparación con sólo un 10 por 100 de los machos que sirven de control en el experimento, cuya charretera se pinta con un disolvente transparente. Las charreteras probablemente mantienen a los intrusos a raya señalando que un territorio está ocupado. El canto también es importante. Los machos enmudecidos, privados temporalmente de su capacidad de cantar, también pierden territorios.
En los turpiales, pues, el canto y el plumaje ayudan al macho a conseguir más parejas. En los estudios descritos anteriormente, y en muchos otros, los investigadores han mostrado que la selección sexual está actuando porque los machos con los rasgos más elaborados obtienen un beneficio mayor en términos de descendencia. Esta conclusión parece simple pero ha precisado cientos de horas de tedioso trabajo de campo por parte de biólogos animados por la curiosidad. La secuenciación de ADN en un reluciente laboratorio puede parecer mucho más glamuroso, pero sólo ensuciándose en el campo puede un científico decirnos cómo opera la selección en la naturaleza.
La selección sexual no se agota en el acto sexual: los machos pueden continuar compitiendo incluso después de aparearse. En muchas especies, las hembras se aparean con más de un macho en un breve período de tiempo. Después de que un macho haya inseminado a una hembra, ¿cómo puede impedir que otros machos la fecunden y le roben la paternidad? Esta competencia postapareamiento ha producido algunos de los rasgos más curiosos que haya fabricado la selección sexual. A veces un macho se queda con la hembra después de aparearse para impedir que accedan a ella otros pretendientes. Cuando vemos un par de libélulas enganchadas, es probable que el macho simplemente esté controlando a la hembra después de fecundarla, bloqueando físicamente el acceso a otros machos. Un milpiés de América Central ha llevado el control de la hembra al extremo: después de fecundarla, el macho monta sobre su cuerpo durante varios días para impedir que algún competidor se lleve sus huevos. También se pueden usar sustancias químicas para conseguir lo mismo. La eyaculación de algunas serpientes y roedores contiene sustancias que de manera temporal obstruyen el tracto reproductor de la hembra después del apareamiento, impidiendo que otros machos puedan copular con ella. En el grupo de las moscas de la fruta con las que investigo, el macho inyecta en la hembra un antiafrodisíaco, una sustancia química que lleva en el semen para quitarle las ganas de aparearse durante varios días.
Los machos utilizan todo tipo de armas defensivas para proteger su paternidad. Pero pueden ser aún más taimados y utilizar armas ofensivas para deshacerse del esperma de los machos que hayan copulado antes que ellos, y reemplazarlo con el propio. Uno de los artilugios más ingeniosos es el «pene en escobilla» de algunos caballitos del diablo. Cuando un macho se aparea con una hembra que ya había sido fecundada, utiliza las cerdas dirigidas hacia atrás de su pene para extraer el esperma de los machos que lo precedieron. Sólo después de haber limpiado a la hembra de esperma le transfiere el suyo propio. Con las moscas del género Drosophila, mi propio laboratorio descubrió que la eyaculación de un macho contiene sustancias que inactivan el esperma almacenado de los machos que hayan copulado antes.
¿Y qué pasa con la segunda forma de selección sexual, la elección de pareja? En comparación con la competencia entre machos, es mucho menos lo que sabemos sobre cómo funciona este proceso. La razón es que el significado de los colores, el plumaje y la exhibición es mucho menos obvio que el de las cornamentas y otras armas.
Para entender cómo ha evolucionado la elección de pareja, comencemos por esa enojosa cola de pavo real que tanta angustia causaba a Darwin. Buena parte de las investigaciones sobre la elección de pareja en el pavo real han sido realizadas por Marion Petrie y sus colegas, que estudiaron una población asilvestrada en Whipsnade Park, en Bedfordshire (Inglaterra). En esta especie los machos se reúnen en leks, los lugares donde todos juntos hacen sus exhibiciones, ofreciendo así a las hembras la oportunidad de compararlos directamente. No todos los machos participan en el lek, pero sólo quienes lo hacen pueden conseguir una hembra. En un estudio observacional de diez machos que participaban en un lek se encontró una fuerte correlación entre el número de ocelos en las plumas de la cola del macho y el número de apareamientos conseguidos: el macho más ornamentado, con 160 ocelos, consiguió el 36 por 100 de todas las cópulas.
Esto sugiere, pero no demuestra, que las colas más ornamentadas son las preferidas por las hembras. Es posible que algún otro aspecto del cortejo del macho, por ejemplo el vigor de su exhibición, sea realmente lo que las hembras eligen, y que este otro aspecto esté correlacionado con el plumaje. Para descartar esta posibilidad, pueden realizarse manipulaciones experimentales: cambiar el número de ocelos en la cola de un pavo real y ver si esto afecta a su capacidad de conseguir parejas. Curiosamente, este mismo experimento fue sugerido ya en 1869 por el competidor de Darwin, Alfred Russel Wallace. Aunque los dos hombres se mostraban de acuerdo en muchas cosas, y en particular en la selección natural, discrepaban sobre la selección sexual. La idea de la competencia entre machos no era un problema para ninguno de los dos, pero Wallace desaprobaba la posibilidad de la elección por las hembras. No obstante, mantuvo una actitud abierta sobre esta cuestión, y se adelantó mucho a su tiempo cuando sugirió cómo contrastarla:
La parte que desempeñaría sólo el ornamento sería muy pequeña, aunque estuviera demostrado, que no lo está, que una ligera superioridad únicamente en el ornamento decida usualmente la elección de pareja.
Esto, sin embargo, es un aspecto que se presta a la experimentación, y me permito sugerir que algún miembro de la Sociedad Zoológica o cualquiera que disponga de los medios, intente realizar tales experimentos. Debería elegirse para ello una docena de machos de la misma edad —de gallos domésticos, faisanes comunes o faisanes dorados, por ejemplo—, todos ellos de condición que se sepa aceptable para las hembras del ave. A la mitad de éstos debería cortárseles una o dos plumas de la cola, o reducir un poco de tamaño las plumas del cuello, apenas lo justo para producir una diferencia que pudiera darse en la naturaleza, pero que no llegue a desfigurar al ave, y luego se debería observar si las hembras notan de algún modo la deficiencia, y si de manera uniforme rechazan a los machos menos ornamentados. Tales experimentos, si se realizan con todo el cuidado y se varían de manera juiciosa durante varias estaciones, arrojarían información muy valiosa sobre esta interesante cuestión.
Estos experimentos no se realizaron hasta más de un siglo después. En cualquier caso, por fin tenemos los resultados: la elección por la hembra es común. En uno de estos experimentos, Marion Petrie y Tim Halliday cortaron veinte ocelos de la cola de cada uno de los machos de un grupo de pavos reales, y compararon su éxito de apareamiento con el de un grupo de control que fue manipulado, pero sin cortarle nada. En la siguiente época de cría los machos con reducción de ornamentos consiguieron por término medio 2,5 apareamientos menos que los machos del grupo de control.
Este experimento sugiere claramente que las hembras prefirieron a los machos cuyos ornamentos no habían sido reducidos. Pero, idealmente, deberíamos hacer el experimento también en el sentido contrario: ornamentar más las colas para ver si de este modo se aumenta el éxito de apareamiento. Esto es difícil de hacer en los pavos reales, pero el biólogo sueco Malte Andersson ha logrado hacerlo con un ave territorial, el obispo de cola larga (Euplectes progne). En esta especie sexualmente dimórfica, los machos tienen colas de unos cincuenta centímetros de longitud y las hembras de sólo siete centímetros. Andersson cortó una parte de la larga cola de algunos machos y algunas de las partes cortadas las pegó a las colas normales de otros machos; de este modo, consiguió tener un grupo de machos de cola corta (15 centímetros), un grupo de control con la cola normal (con la cola cortada y vuelta a pegar) y un grupo de cola larga (75 centímetros). Tal como se esperaba, los machos de cola corta consiguieron atraer a menos hembras para que nidificasen en su territorio por comparación con los machos normales. Y los machos con la cola artificialmente larga obtuvieron un aumento fenomenal del número de apareamientos, atrayendo casi el doble de hembras que los machos normales.
Esto plantea una pregunta: si los machos con una cola de 75 centímetros consiguieron más hembras, ¿por qué los obispos no habían evolucionado ya hasta obtener una cola tan larga? No sabemos la respuesta, pero es probable que una cola tan larga reduzca la longevidad del macho más de lo que aumenta su capacidad para conseguir parejas. Cincuenta centímetros probablemente sea la longitud para la cual se obtenga, por término medio, el mayor número de descendientes en el curso de la vida reproductora.
¿Y qué consiguen los machos del gallo de las artemisas con sus arduas payasadas en la pradera? Una vez más, la respuesta es más apareamientos. Como los pavos reales, los machos del gallo de las artemisas forman leks para exhibirse en masa mientras las hembras pasan revista. Se ha podido mostrar que sólo los machos más vigorosos, los que «aletean» alrededor de ochocientas veces al día, consiguen hembras, y que la gran mayoría de los machos se quedan sin aparearse.
La selección sexual también explica las proezas arquitectónicas de las aves de emparrado. Varios estudios han mostrado que los tipos de emparrado, que difieren entre especies, están correlacionados con el éxito en el apareamiento. Por ejemplo, en el capulinero satinado, los machos consiguen más hembras si ponen más plumas azules en sus emparrados. En el capulinero moteado, el mayor éxito se obtiene adornándolos con bayas verdes de Solanum (una especie de tomate silvestre). Joah Madden, de la Universidad de Cambridge, quitó las decoraciones de los emparrados de capulineros moteados y luego ofreció a los machos sesenta objetos entre los que elegir. Indefectiblemente, volvieron a decorar sus emparrados sobre todo con bayas de Solanum, que colocaron en los lugares más visibles.
Me he centrado en las aves porque a los biólogos les ha resultado más fácil estudiar la elección de pareja en este grupo debido a que las aves son activas durante el día y fáciles de observar, pero hay muchos ejemplos de elección de pareja en otros animales. Las hembras de rana túngara prefieren aparearse con machos que croan los cantos más complejos, las hembras de gupi prefieren los machos con la cola más larga y manchas de colores más vivos, y las hembras de las arañas y de los peces suelen sentir preferencia por los machos de mayor tamaño. En su exhaustiva obra Sexual Selection, Malte Andersson describe 232 experimentos con 186 especies que muestran que una gran variedad de rasgos de los machos están correlacionados con el éxito en el apareamiento, y la mayoría de estos ensayos implican la elección por las hembras. Sencillamente, no hay ninguna duda de que la elección por las hembras ha impulsado la evolución de muchos dimorfismos sexuales. Al final, era Darwin quien tenía razón.
Hasta este momento hemos desatendido dos cuestiones importantes: ¿por qué son las hembras quienes eligen mientras los machos las cortejan o luchan por ellas? Y ¿por qué elegir siquiera? Para responder a estas dos preguntas primero tenemos que entender por qué los organismos se molestan en tener sexo.
¿Por qué el sexo?
Porqué evolucionó el sexo es, de hecho, uno de los mayores misterios de la evolución. Todo individuo que se reproduce sexualmente (es decir, fabricando óvulos o espermatozoides que contienen la mitad de sus genes) sacrifica el 50 por 100 de su contribución genética a la siguiente generación en comparación con un individuo que se reproduzca asexualmente. Supongamos que existiera en los humanos un gen cuya forma normal condujera a la reproducción sexual mientras que su forma mutante permitiera a las hembras reproducirse partenogenéticamente, es decir, produciendo huevos que se desarrollen sin ser fecundados. (Algunos animales se reproducen de este modo: se ha visto, por ejemplo, en pulgones, peces y lagartos.) La primera mujer mutante sólo tendría hijas, que a su vez tendrían más hijas. En cambio, las mujeres no mutantes, con reproducción sexual, tendrían que aparearse con machos, y por término medio tendrían una mitad de hijos y la otra mitad de hijas. La proporción de mujeres en la población aumentaría rápidamente por encima del 50 por 100 a medida que el conjunto de las mujeres se fuera llenando de mutantes producidas asexualmente por madres de reproducción asexual. Los machos acabarían siendo superfluos y desaparecerían: ninguna mujer mutante necesitaría aparearse con ellos, y todas las mujeres tendrían solamente hijas. El gen de la partenogénesis habría ganado la competición contra el gen de la reproducción sexual, desplazándolo. Puede demostrarse teóricamente que en cada generación los genes «asexuales» producirían el doble de copias de sí mismos que el gen «sexual» original. Los biólogos denominan a esta situación «el doble coste del sexo». El resultado final es que bajo la selección natural los genes de la partenogénesis se extienden con rapidez, eliminando la reproducción sexual.
Pero no es esto lo que ha ocurrido en la realidad. La gran mayoría de las especies de la Tierra se reproducen sexualmente, y esta forma de reproducción se viene haciendo desde hace más de 1.000 millones de años.[36] ¿Por qué el coste del sexo no ha conducido a que sea reemplazado por la partenogénesis? Está claro que el sexo debe ofrecer alguna ventaja evolutiva que compense con creces su coste. Aunque todavía no sabemos con certeza cuál es esa ventaja, teorías no nos faltan.
La clave podría estar en la mezcla al azar de genes que se produce durante la reproducción sexual, que genera nuevas combinaciones de genes en la descendencia. Al juntar varios genes favorables en un mismo individuo, el sexo podría promover una evolución más rápida y una mejor respuesta a aspectos del entorno que cambien continuamente, como los parásitos, que evolucionan sin tregua para contrarrestar la evolución de nuestras propias defensas. O quizá el sexo purgue los genes malos de una especie al recombinarlos en un individuo con graves desventajas, una suerte de cabeza de turco genética. Pero los biólogos todavía se cuestionan si alguna de las ventajas conocidas compensa con creces el doble coste del sexo.
Una vez el sexo ha evolucionado, la selección sexual se sigue de forma ineludible siempre y cuando podamos explicar dos cosas más. La primera es por qué hay sólo dos sexos (y no tres o más) que tengan que combinar sus genes para producir descendencia. La segunda es por qué los dos sexos tienen un número desigual de gametos de diferente tamaño (los machos producen un montón de espermatozoides mientras que las hembras producen un número menor de óvulos de mayor tamaño). La pregunta del número de sexos es una compleja cuestión teórica que no debe detenernos, salvo para hacer notar que la teoría muestra que un sistema con dos sexos reemplaza evolutivamente a un sistema con tres o más sexos, es decir, dos sexos es la estrategia estable más robusta.
La teoría de por qué los dos sexos tienen gametos que difieren en número y en tamaño es igualmente compleja. Esta condición supuestamente evolucionó a partir de una más primitiva en la que las especies de reproducción sexual producían gametos de igual tamaño. Los teóricos han demostrado de manera bastante convincente que la selección natural favorece el cambio de este estado ancestral a un estado en el que un sexo (el que llamamos «macho») produce muchos gametos pequeños (espermatozoides o polen) y el otro (la «hembra») produce menos gametos, pero más grandes (los óvulos o huevos).
Es esta asimetría en el tamaño de los gametos lo que prepara el terreno para toda la selección sexual, pues hace que los dos sexos evolucionen hacia estrategias de apareamiento distintas. Centrémonos en el macho. Un macho puede producir grandes cantidades de espermatozoides, y por lo tanto en principio podría ser el padre de un gran número de descendientes, limitado únicamente por el número de hembras que pueda atraer y por la capacidad competitiva de sus espermatozoides. Para las hembras, en cambio, las cosas son distintas. Los óvulos son costosos y están en número limitado, y si una hembra se aparea muchas veces en un corto período de tiempo, es poco (o nada) lo que hace para aumentar el número de descendientes.
Una demostración muy vistosa de esta diferencia se puede ver en el número récord de hijos de un hombre o de una mujer. Si el lector hubiera de conjeturar cuál es el número máximo de hijos que puede tener una mujer durante toda su vida, seguramente diría en torno a quince. Mejor que pruebe otra vez. El Libro Guinness de los récords, nos dice que el récord «oficial» de hijos de una mujer es de sesenta y nueve, y lo ostenta una campesina rusa del siglo XIX que en los veintisiete embarazos que tuvo entre 1725 y 1745, alumbró mellizos dieciséis veces, trillizos siete veces y cuatrillizos cuatro veces. (Presuntamente tenía alguna predisposición fisiológica o genética a los embarazos múltiples.) Uno compadece a esta esforzada mujer, pero su récord es superado en mucho por el récord de un hombre, un tal Mulai Ismail (1646-1727), emperador de Marruecos. Ismail fue padre, según nos dice el Guinness, de «al menos 342 hijas y 525 hijos, y se dice que en 1721 tenía 700 descendientes varones». Incluso en estos extremos, los machos superan a las hembras con una descendencia diez veces mayor.
La diferencia evolutiva entre machos y hembras es una cuestión de inversión diferencial: inversión en huevos caros frente a espermatozoides baratos, inversión en el embarazo (cuando las hembras retienen y nutren los huevos fecundados), e inversión en el cuidado parental en las muchas especies en las que la hembra es la única que cría a los jóvenes. Para los machos, aparearse es barato; para las hembras es caro. Para los machos, el apareamiento sólo cuesta una pequeña dosis de esperma; para las hembras, cuesta mucho más: la producción de óvulos grandes y ricos en nutrientes y, con frecuencia, un enorme gasto de tiempo y energía. En más del 90 por 100 de las especies de mamíferos, la única inversión del macho en la descendencia es el esperma, pues son las hembras las que proporcionan el cuidado parental.
Esta asimetría entre machos y hembras en el número potencial de apareamientos y descendientes conduce a conflictos de intereses en el momento de escoger una pareja. Los machos tienen poco que perder apareándose con una hembra «por debajo de la media» (por ejemplo, una que sea débil o esté enferma), porque no les cuesta casi nada aparearse otra vez, y así las veces que haga falta. Por tanto, la selección favorecerá los genes que hagan machos promiscuos que intenten aparearse con todas las hembras que puedan. (O con cualquier cosa que se parezca en lo más mínimo a una hembra; el gallo de las artemisas, por ejemplo, a veces intenta copular con pilas de estiércol de vaca, y, como ya hemos visto, algunas orquídeas consiguen polinizarse atrayendo a machos calientes de abeja que intentan copular con sus pétalos.)
Las hembras son distintas. A causa de su mayor inversión en huevos y descendientes, su mejor táctica consiste en ser exigentes en lugar de promiscuas. Las hembras tienen que conseguir que cada oportunidad cuente eligiendo al mejor padre posible para fecundar su limitado número de huevos. Por eso tienen que inspeccionar muy de cerca a sus pretendientes.
El resultado de todo esto es que, por lo general, los machos tienen que competir por las hembras. Los machos deberían ser promiscuos, las hembras recatadas. La vida de un macho debería ser de conflicto constante con sus iguales, siempre compitiendo con los otros machos por las parejas. Los buenos machos, más atractivos o más vigorosos, se llevarán siempre un gran número de parejas (presuntamente serán preferidos también por más hembras), mientras que los inferiores se quedarán sin aparearse. Casi todas las hembras, en cambio, acabarán por encontrar pareja. Como todos los machos compiten por ellas, su distribución de éxito de apareamiento será más uniforme.
Los biólogos describen esta diferencia diciendo que la varianza del éxito de apareamiento debería ser mayor para los machos que para las hembras. ¿Ocurre así? En efecto, a menudo observamos esta diferencia. En el ciervo común, por ejemplo, la variación en el número de descendientes producidos durante toda la vida es tres veces mayor entre machos que entre hembras. La disparidad es aún mayor para los elefantes marinos, en los que menos del 10 por 100 de los machos dejan algún descendiente a lo largo de varias épocas de cría, en comparación con más de la mitad de las hembras.[37]
La diferencia entre machos y hembras en el número potencial de descendientes impulsa la evolución tanto de la competencia entre machos como de la elección por las hembras. Los machos tienen que competir para fecundar un número limitado de huevos. Por eso vemos la «ley de la batalla»: la competencia directa entre machos para dejar sus genes a la siguiente generación. Y ésa es también la razón de que los machos sean vistosos y coloridos, de que realicen exhibiciones, o emitan canciones de apareamiento, o construyan emparrados u otras cosas por el estilo, pues ésa es su forma de decir «¡escógeme!, ¡escógeme!». En último término es la preferencia de las hembras lo que impulsa la evolución de las colas largas, de las exhibiciones más vigorosas o de las canciones más sonoras en los machos.
Naturalmente, lo que acabo de describir es una generalización: hay excepciones. Algunas especies son monógamas, y tanto el macho como la hembra realizan los cuidados parentales. La evolución puede favorecer la monogamia si los machos tienen más descendientes ayudando a cuidar a las crías que si abandonan a su descendencia para buscar otras parejas. En muchas aves, por ejemplo, se necesitan los dos progenitores a tiempo completo: cuando uno sale a aprovisionarse, el otro incuba los huevos. Pero las especies monógamas no son muy comunes en la naturaleza. Sólo un 2 por 100 de todas las especies de mamíferos, por ejemplo, sigue este sistema de apareamiento.
Además, hay otras explicaciones del dimorfismo sexual en el tamaño corporal en las que no interviene la selección sexual. En las moscas de la fruta que investigo, por ejemplo, las hembras pueden ser mayores sencillamente porque necesitan producir huevos grandes y costosos. O los machos y las hembras podrían ser depredadores más eficientes si se especializan en presas distintas. La selección natural en el sentido de una reducción de la competencia entre los dos sexos puede llevarlos a evolucionar hacia tamaños corporales distintos. Esto explica el dimorfismo en algunos lagartos y aves rapaces, en los que las hembras son mayores que los machos y capturan presas más grandes.
Romper las reglas
Curiosamente, también encontramos casos de dimorfismo sexual en muchas especies «socialmente monógamas», es decir, en las que machos y hembras se emparejan y crían a los jóvenes juntos. Dado que los machos no parecen estar compitiendo por las hembras, ¿por qué han evolucionado hacia colores vivos y ornamentos? Esta aparente contradicción en realidad proporciona apoyo adicional a la teoría de la selección sexual. Resulta que en estos casos las apariencias engañan. Estas especies son socialmente monógamas, pero no realmente monógamas.
Una de estas especies es el maluro espléndido de Australia, un ave que ha estudiado mi colega de la Universidad de Chicago, Stephen Pruett-Jones. A primera vista, esta especie parece un dechado de monogamia. Machos y hembras suelen pasar toda su vida adulta unidos por un vínculo social, marcan su territorio y comparten el cuidado parental. Sin embargo, presentan un sorprendente dimorfismo sexual en el plumaje: los machos son de un hermoso color azul y negro iridiscentes, mientras que las hembras son de un soso color gris pardo. ¿Por qué? Porque el adulterio abunda. Cuando llega el momento de aparearse, las hembras se aparean con otros machos más a menudo que con su «pareja social». (Esto lo sabemos gracias a análisis de paternidad con el ADN.) Los machos juegan al mismo juego, buscando y solicitando apareamientos «extrapareja», pero aun así varían mucho más que las hembras en su éxito reproductor. La selección sexual asociada a estas parejas adultas casi con certeza condujo a la evolución de las diferencias de color entre los dos sexos. Este pájaro no es único en su comportamiento. Aunque el 90 por 100 de las especies de ave son socialmente monógamas, en las tres cuartas partes los machos y las hembras se aparean con individuos distintos de su compañero social.
La teoría de la selección sexual produce predicciones contrastables. Si sólo un sexo tiene plumaje brillante o cornamenta, realiza enérgicas exhibiciones de cortejo o construye complejas estructuras para atraer a las hembras, puede apostarse que son los miembros de ese sexo quienes compiten por aparearse con los miembros del otro. Y las especies que presentan menos dimorfismo sexual en la conducta o la apariencia deberían ser más monógamas: si los machos y las hembras se aparean y se quedan con sus parejas, no hay competencia sexual y no hay, por tanto, selección sexual. De hecho, los biólogos observan una fuerte correlación entre los sistemas de apareamiento y el dimorfismo sexual. Los dimorfismos extremos en el tamaño, el color o el comportamiento se encuentran en las especies en las que, como ocurre con las aves del paraíso o los elefantes marinos, los machos tienen que competir por las hembras, y sólo unos pocos consiguen aparearse. Las especies en las que los machos y las hembras tienen aspecto muy parecido, como los gansos, los pingüinos y los loros, tienden a ser monógamas auténticas, ejemplos de fidelidad animal. Esta correlación es otro triunfo de la teoría de la evolución, pues sólo es predicha por la idea de la selección sexual, y no por ninguna alternativa creacionista. ¿Por qué habría de haber una correlación entre el color y el sistema de apareamiento a no ser que la evolución sea cierta? En realidad, son los creacionistas, y no los evolucionistas, quienes deberían marearse a la vista de una pluma de pavo real.[38]
Hasta el momento hemos hablado de la selección sexual como si el sexo promiscuo fuese siempre el macho y el que escoge fuese la hembra. Sin embargo, en algunas raras ocasiones, ocurre justo al revés. Y cuando estas conductas se intercambian entre los sexos, también lo hace la dirección del dimorfismo. Vemos esta inversión en esos atractivos peces, los caballitos de mar, y en su pariente cercano el pez aguja. En algunas de estas especies no es la hembra la que pare las crías, ¡sino el macho! ¿Cómo puede ser? Aunque la hembra produce los huevos, después de fecundarlos el macho los coloca en el interior de una bolsa incubadora especializada situada en el vientre o en la cola, y los lleva consigo hasta que eclosionan. Los machos sólo llevan cada vez las crías de una hembra, y su período de «gestación» dura más tiempo del que necesita la hembra para producir huevos otra vez. Así que los machos invierten más en las crías que las hembras. Además, como hay más hembras con huevos sin fecundar que machos libres para aceptarlos, las hembras tienen que competir por los raros machos que no están «preñados». En este caso, la diferencia de estrategia reproductora entre machos y hembras queda invertida, y tal como puede esperarse de la teoría de la selección sexual, son las hembras las que están decoradas con colores vivos y ornamentos corporales, mientras que los machos son relativamente insulsos.
Lo mismo puede decirse de los falaropos, tres especies de gráciles aves limícolas que crían en Europa y América del Norte y son de los pocos ejemplos de sistema de apareamiento poliándrico («una hembra y muchos machos»). (Este raro sistema de apareamiento también se da en algunas poblaciones humanas, como los tibetanos.) Los machos de falaropo se ocupan por entero de cuidar las crías, de construir el nido y de alimentar a la pollada mientras la hembra se dedica a buscar otros machos con los que aparearse. La inversión de los machos en la descendencia es, por tanto, mayor que la de las hembras, y las hembras compiten por los machos que habrán de cuidar a sus crías. Como es de esperar, en las tres especies las hembras tienen una coloración más viva que los machos.
Caballitos de mar, peces aguja y falaropos son las excepciones que confirman la regla. Su decoración «invertida» es exactamente lo que uno esperaría encontrar si la explicación evolutiva del dimorfismo sexual fuera cierta, mientras que no tiene ningún sentido si estas especies hubieran sido especialmente creadas.
¿Por qué elegir?
Volvamos ahora a la elección de pareja «normal», aquella en la que son las hembras quienes eligen. ¿Qué es exactamente lo que buscan en el momento de elegir un macho? Esta pregunta ha inspirado un célebre desacuerdo en la biología evolutiva. Alfred Russel Wallace, como ya hemos visto, dudaba incluso de que las hembras eligieran (pero, como sabemos, se equivocaba). Su propia teoría era que las hembras tenían menos colorido que los machos porque necesitaban camuflarse de los depredadores, mientras que los colores vivos y los ornamentos de los machos eran productos secundarios de su fisiología. No ofrecía ninguna explicación, sin embargo, de por qué los machos no deberían camuflarse también.
La teoría de Darwin era algo mejor. Creía con firmeza que los cantos, colores y ornamentos de los machos habían evolucionado en respuesta a la elección por las hembras. Pero ¿a qué atendían las hembras al elegir? Su respuesta fue sorprendente: pura estética. Darwin no veía razón alguna para que las hembras eligieran cosas como un canto complejo o una cola larga a no ser que les resultaran intrínsecamente atractivas. Su obra pionera sobre la selección sexual, El origen del hombre y la selección con relación al sexo (1871), está cargada de pintorescas descripciones antropomórficas de cómo las hembras de los animales se sienten «cautivadas» o «atraídas» por diversas características de los machos. Sin embargo, como Wallace bien observó, todavía quedaba un problema. ¿Tenían realmente los animales, y sobre todo los más simples, como los escarabajos y las moscas, un sentido estético como el nuestro? Darwin salió del paso como pudo, alegando ignorancia:
Aunque tenemos indicios positivos de que las aves aprecian los objetos bellos y brillantes, como ocurre con las aves de emparrado de Australia, y aunque no cabe duda de que aprecian el poder del canto, debo admitir sin ambages que es sorprendente que las hembras de muchas aves y algunos mamíferos estén dotadas del gusto suficiente para apreciar los ornamentos que tenemos razones para atribuir a la selección sexual; y esto es todavía más sorprendente en el caso de los reptiles, los peces y los insectos. Pero es realmente poco lo que sabemos de la mente de los animales inferiores.
Aunque no tenía todas las respuestas, hoy sabemos que Darwin estaba más cerca de la verdad que Wallace. Sí, las hembras eligen, y esa elección parece explicar el dimorfismo sexual. Pero no tiene sentido que la preferencia de las hembras se base únicamente en la estética. Especies estrechamente emparentadas, como las aves del paraíso de Nueva Guinea, tienen machos con tipos de plumaje y conductas de apareamiento muy distintos. ¿Tan diferente es lo que es hermoso para una especie de lo que lo es para sus parientes más cercanos?
En realidad, en la actualidad tenemos muchos y buenos indicios de que las preferencias de las hembras son en sí mismas adaptativas, porque preferir ciertos tipos de macho ayuda a las hembras a diseminar sus genes. Las preferencias no son siempre una cuestión de gusto congénito y aleatorio, como Darwin suponía, sino que en muchos casos probablemente evolucionaron por selección natural.
¿Qué es lo que puede ganar una hembra al seleccionar a un macho determinado? Hay dos respuestas a esta pregunta. Puede beneficiarse directamente, es decir, eligiendo un macho que la ayude a producir más crías o crías más sanas durante el acto del cuidado parental. O puede beneficiarse indirectamente escogiendo un macho que tenga mejores genes que otros machos (es decir, genes que den a su descendencia una ventaja en la siguiente generación). En cualquier caso, la evolución de las preferencias de las hembras se verá favorecida por la selección. Por la selección natural.
Veamos primero el caso de los beneficios directos. Un gen que le diga a una hembra que se aparee con machos que guarden un territorio mejor le proporcionará descendientes que estarán más nutridos o que ocuparán mejores nidos, lo que significa que sobrevivirán mejor y se reproducirán más que las crías que no hayan crecido en territorios de la misma calidad. Esto implica que la población de las crías llevará una proporción de hembras con el «gen de la preferencia» mayor que la generación anterior. A medida que pasen las generaciones y prosiga la evolución, todas las hembras llevarán los genes de la preferencia. Y si apareciera otra mutación que incrementara la preferencia por territorios mejores, también ésta aumentará en frecuencia. Con el tiempo, la preferencia por los machos con mejores territorios irá evolucionando y haciéndose cada vez más fuerte. Esto, a su vez, seleccionará a los machos que compitan mejor por los territorios. La preferencia de las hembras evoluciona en paralelo a la competencia entre los machos por los territorios.
Los genes que reportan beneficios indirectos a las hembras selectivas también se extenderán. Imaginemos que un macho tiene genes que lo hacen más resistente a las enfermedades que otros machos. Una hembra que se aparee con uno de estos machos tendrá descendencia más resistente a las enfermedades. Esto le otorga un beneficio evolutivo por haber elegido ese macho. Ahora imaginemos que además existe un gen que permite a las hembras identificar como pareja a los machos más sanos. Si una hembra se aparea con unos de esos machos, el resultado será un conjunto de hijos e hijas con ambos tipos de genes, los que otorgan resistencia a las enfermedades y los que llevan a preferir a los machos con resistencia a la enfermedad. En cada generación, los individuos más resistentes a las enfermedades, que se reproducirán con más éxito, también llevarán los genes que les dicen a las hembras que escojan a los machos más resistentes. Los genes de la resistencia se extenderán por selección natural, y los genes de la preferencia de los machos los acompañarán. De este modo aumentarán en la especie tanto la preferencia de las hembras como la resistencia a las enfermedades.
Estas dos explicaciones hipotéticas explican por qué las hembras prefieren ciertos tipos de machos, pero no por qué prefieren ciertos caracteres de esos machos, como los colores vivos o los elaborados plumajes. Esto probablemente pasa porque esos caracteres en particular le dicen a la hembra que un macho le proporcionará unos mayores beneficios directos o indirectos.
El pinzón mexicano (Carpodacus mexicanus) de América del Norte es sexualmente dimórfico respecto al color: las hembras son pardas pero los machos tienen colores vivos en la cabeza y el pecho. Los machos no defienden un territorio pero participan en el cuidado parental. Geoff Hill, de la Universidad de Michigan, encontró que en una población local los machos variaban en color desde el amarillo pálido, pasando por el naranja, hasta el rojo intenso. Para saber si el color afectaba al éxito reproductor, utilizó tintes para hacer a los machos más brillantes o más pálidos. Y, en efecto, halló que los machos de colores más intensos obtenían una cantidad significativamente mayor de parejas que los pálidos. Además, entre los pájaros no manipulados, las hembras abandonaron los nidos de los machos más pálidos más a menudo que los de los machos de colores más vivos.
¿Por qué las hembras de pinzón mexicano prefieren los machos de colores más vivos? Hill mostró que en la misma población los machos de más color alimentaban a sus pollos más a menudo que los machos más pálidos. Así que las hembras obtenían un beneficio directo, en forma de un mejor aprovisionamiento para su pollada, cuando elegían un macho de colores más vivos. (Las hembras que se apareaban con machos más pálidos podían llegar a abandonar el nido porque sus crías no recibían la alimentación adecuada.) Pero ¿por qué traen más comida los machos de colores más vivos? Probablemente lo que ocurre es que la viveza del color es un signo de su salud general. El color rojo de estos machos proviene en su totalidad de los pigmentos carotenoides de las semillas que comen: no pueden sintetizarlo ellos mismos. Los machos de colores más vivos están, por lo tanto, mejor alimentados, y probablemente gocen de mejor salud general. Al parecer, las hembras escogen a los machos de color más intenso porque el color les dice «soy un macho más capaz de llenar la despensa de la familia». Cualquier gen que haga que las hembras prefieran los machos de colores vivos les reportará un beneficio directo, de manera que la selección aumentará esa preferencia. Una vez establecida la preferencia, cualquier macho con una mayor capacidad para convertir las semillas en plumaje de color vivo también obtendrá una ventaja, ya que conseguirá más parejas. Con el tiempo, la selección sexual exagerará el color rojo del macho. Las hembras mantienen el color pálido porque un color vivo no les reporta ningún beneficio, y en cambio las hace más visibles a los depredadores.
Escoger un macho fuerte y sano conlleva también otros beneficios. Los machos pueden llevar parásitos o enfermedades que pueden transmitir a las hembras, a las crías o a ambas, así que beneficia a las hembras evitar esos machos. El color, el plumaje y el comportamiento de un macho pueden servir de pista para saber si está enfermo o infestado: sólo los machos sanos pueden cantar con voz potente, realizar una exhibición vigorosa o producir una serie de plumas brillantes y elegantes. Si, por poner un caso, los machos de una especie son normalmente de color azul intenso, convendrá evitar aparearse con un macho de color azul pálido.
La teoría de la evolución indica que las hembras deberían preferir cualquier rasgo que muestre que un macho puede ser un buen padre. Basta con que haya algún gen que aumente la preferencia por ese rasgo, y que la variación en la expresión del rasgo sea una buena indicación de la condición física del macho. El resto se sigue de manera automática. En el gallo de las artemisas, unos piojos parásitos producen unos puntos de sangre en el saco vocal del macho que son muy visibles cuando, durante la exhibición de aleteo en el lek, el macho hincha el saco hasta el punto de ser translúcido. Los machos a los que se pinta unos falsos puntos de sangre en los sacos vocales obtienen un número significativamente menor de apareamientos, lo que sugiere que los puntos indican a las hembras que ese macho está infestado y que, literalmente, como padre sería un piojoso. La selección favorecerá los genes que promuevan no sólo la preferencia de las hembras por los sacos sin puntos, sino también el carácter que en los machos indique este problema. El saco vocal del macho se hará mayor, y la preferencia de la hembra por el saco vocal sin manchas aumentará. Esto puede conducir a la evolución de caracteres muy exagerados en los machos, como la cola ridículamente larga de los obispos. El proceso de exageración del carácter del macho sólo se frena cuando se llega al punto en que cualquier aumento adicional reduce la supervivencia del macho más de lo que atrae a las hembras y la producción neta de descendientes del macho se resiente.
¿Qué ocurre, sin embargo, con las preferencias de las hembras que proporcionan beneficios indirectos? El más obvio de estos beneficios es lo que un macho siempre le da a sus descendientes: sus genes. El mismo tipo de caracteres que indican que un macho es sano podrían mostrar también que está genéticamente bien dotado. Quizá los machos con colores más vivos, colas más largas o cantos más potentes puedan exhibir esos rasgos sólo si poseen los genes que los hacen sobrevivir más o reproducirse mejor que sus competidores. Lo mismo puede decirse de los machos capaces de construir complejos emparrados, o de apilar grandes mojones de piedras. Podemos imaginar muchos caracteres que podrían mostrar que un macho tiene genes que le otorgan una mayor supervivencia o una mayor capacidad reproductora. La teoría evolutiva nos dice que en estos casos habrá tres tipos de genes que aumentarán su frecuencia conjuntamente: los genes de un carácter «indicador» del macho que refleje que posee buenos genes, los genes que hacen que una hembra prefiera un carácter indicador, y, naturalmente, los genes «buenos» cuya presencia refleja el carácter indicador. Es una hipótesis compleja, pero la mayoría de los biólogos evolutivos creen que es la mejor explicación de los caracteres y conductas complejos de los machos.
Pero ¿cómo podemos poner a prueba el modelo de los «genes buenos»? ¿Buscan las hembras los beneficios directos o los indirectos? Una hembra puede rechazar a un macho menos vigoroso o menos vistoso, pero esto podría reflejar no tanto su pobre dotación genética como, sencillamente, una debilidad con causa ambiental, por ejemplo una infección o malnutrición. Estas complicaciones hacen que las causas de la selección sexual sean realmente difíciles de desentrañar en cada caso particular.
Quizá el mejor experimento que pone a prueba el modelo de los genes buenos sea el que realizaron Allison Welch, y sus colegas de la Universidad de Misuri, con la rana arbórea gris (Hyla versicolor). Los machos de esta rana atraen a las hembras con unos sonoros cantos que embelesan en las noches del sur de Estados Unidos. Los estudios realizados con ranas cautivas muestran que las hembras manifiestan una fuerte preferencia por los machos con canciones más largas. Para contrastar si esos machos tienen mejores genes, los investigadores sacaron los huevos de varias hembras y fecundaron la mitad de ellos con esperma de machos de canto largo, y la otra mitad con esperma de machos de canto corto. Los renacuajos nacidos de estos cruces se criaron en cautividad hasta alcanzar la madurez. Los resultados fueron espectaculares. La descendencia de los machos con cantos más largos crecieron más rápido y sobrevivieron mejor el estadio de renacuajo, presentaron mayor tamaño en el momento de la metamorfosis (cuando los renacuajos se convierten en ranas) y crecieron más rápido después de la metamorfosis. Como los machos de las ranas arbóreas grises no aportan a la descendencia más que sus genes, las hembras no pueden obtener beneficios directos por escoger un macho que realice un canto largo. Este experimento sugiere de un modo sólido que un canto largo es señal de un macho sano con buenos genes, y que las hembras que escogen esos machos producen una descendencia genéticamente superior.
¿Y los pavos reales? Hemos visto que las hembras prefieren aparearse con los machos que tienen más ocelos en la cola. Además, los machos no participan en la cría de los pollos. Durante sus investigaciones en Whipsnade Park, Marion Petrie pudo mostrar que los machos que tienen más ocelos producen descendientes que crecen más rápido y sobreviven mejor. Es probable que al escoger las colas más adornadas, las hembras escojan los mejores genes, pues son los machos físicamente bien dotados los que pueden producir una cola más elaborada.
Estos dos estudios son toda la evidencia que tenemos hasta el momento de que las hembras escogen a los machos que tienen los mejores genes. Un considerable número de estudios no han encontrado ninguna asociación entre las preferencias de apareamiento y la calidad genética de la descendencia. Pese a ello, el modelo de los buenos genes sigue siendo la explicación preferida de la selección sexual. Que con tan pocos indicios se sostenga esta creencia podría en parte reflejar la preferencia de los evolucionistas por las explicaciones darwinistas estrictas, la creencia en que, de algún modo, las hembras deben poder discriminar entre los genes de los machos.
Existe, sin embargo, una tercera explicación de los dimorfismos sexuales, la más simple de todas. Se fundamenta en los llamados modelos de sesgo sensorial. Estos modelos suponen que la evolución de los dimorfismos sexuales viene impulsada sencillamente por sesgos preexistentes en el sistema nervioso de las hembras. Estos sesgos podrían ser un producto secundario de la selección natural de alguna función diferente de la elección de pareja, por ejemplo la de buscar alimento. Pongamos por caso que los miembros de una especie han desarrollado por medio de la evolución una preferencia visual por el color rojo porque esa preferencia los ayuda a localizar bayas y frutos rojos y maduros. Si apareciera un macho mutante con una mancha de color rojo en el pecho, quizá las hembras lo preferirían sólo porque ya tienen una predilección por el color rojo. En este caso, los machos rojos tendrían una ventaja, y se produciría la evolución de un dimorfismo. (Suponemos que el color rojo representa una desventaja para las hembras porque atrae a los depredadores.) Alternativamente, quizá las hembras sencillamente tienen preferencia por los caracteres nuevos que de algún modo estimulan su sistema nervioso. Podrían, por ejemplo, preferir machos más grandes, machos que retengan su interés con exhibiciones más complejas o machos con una forma rara porque tienen un cola larga. A diferencia de los modelos descritos anteriormente, en el modelo del sesgo sensorial las hembras no obtienen ningún beneficio directo o indirecto por escoger uno u otro macho.
Para poner a prueba esta teoría se puede producir un carácter nuevo en los machos para ver si a las hembras les gusta. Esto es lo que hicieron Nancy Burley y Richard Symanski, de la Universidad de California, con dos especies de pinzones australianos. Se limitaron a pegar a la cabeza de los machos una única pluma en posición vertical, formando una cresta artificial, y luego presentaron a las hembras estos machos crestados y otros sin cresta. (Los pinzones australianos no tienen cresta, pero sí otras especies emparentadas, las cacatúas.) El resultado fue que las hembras mostraron una fuerte preferencia por los machos con cresta artificial de color blanco por comparación con los machos con cresta de color rojo o verde, o los machos normales no crestados. No sabemos por qué las hembras prefieren el blanco, pero quizá sea porque revisten su nido con plumas blancas para camuflar los huevos de la vista de los depredadores. Experimentos similares con ranas y peces muestran asimismo que las hembras tienen preferencia por caracteres a los que nunca habían estado expuestas.[39] El modelo del sesgo sensorial podría ser importante porque la selección natural puede crear a menudo preferencias preexistentes que ayuden a los animales a sobrevivir y reproducirse, y estas preferencias pueden ser aprovechadas por la selección natural para crear nuevos caracteres en los machos. Quizá la teoría de la estética animal de Darwin fuese parcialmente correcta, aunque antropomorfizara las preferencias de las hembras como un «gusto por la belleza».
El lector habrá echado en falta en este capítulo cualquier discusión sobre nuestra propia especie. ¿Qué pasa con nosotros? Hasta qué punto las teorías de la selección sexual se aplican a los humanos es una cuestión compleja que abordaremos en el capítulo 9.