Capítulo 4
Siendo naturalista a bordo del HMS Beagle, me impresionaron mucho ciertos hechos de la distribución geográfica de los habitantes de América del Sur, y de las relaciones geológicas entre los habitantes actuales y los pasados de aquel continente. Estos hechos parecían dar alguna luz sobre el origen de las especies, este misterio de los misterios, como lo ha llamado uno de nuestros mayores filósofos.
CHARLES DARWIN, El origen de las especies
Uno de los lugares más solitarios de la Tierra son las lejanas islas volcánicas de los mares del sur. En una de ellas, Santa Elena, situada a medio camino entre África y América del Sur, transcurrieron los últimos cinco años del cautiverio británico de Napoleón, exiliado de su Francia nativa. Pero las islas más famosas por su aislamiento son las del archipiélago de Juan Fernández, cuatro pequeños retazos de tierra con una superficie total de unos cien kilómetros cuadrados situados a unos 600 kilómetros al oeste de Chile. Fue en una de estas islas donde transcurrió la solitaria vida de náufrago de Alexander Selkirk, el auténtico Robinson Crusoe.
Nacido en 1676 como Alexander Selcraig, Selkirk fue un iracundo escocés que se embarcó en 1703 como maestre en el Cinque Ports, un buque con patente de corso del Gobierno británico para saquear los barcos españoles y portugueses. Preocupado por la temeridad de su joven capitán de veintiún años y por la deplorable condición en que se hallaba el buque, Selkirk exigió que se le dejara en tierra, a la espera de un pronto rescate, cuando el Cinque Ports lanzó amarras para aprovisionarse de agua y alimentos en la isla de Más a Tierra, en el archipiélago de Juan Fernández. El capitán accedió, y Selkirk fue abandonado voluntariamente, llevándose consigo a la isla únicamente ropa, sábanas y algunas herramientas, un fusil de pedernal, tabaco, una tetera y una Biblia. Así comenzaron cuatro años y medio de soledad.
Más a Tierra era una isla desierta en la que los únicos mamíferos, aparte de Selkirk, eran cabras, ratas y gatos que otros navegantes habían introducido en el pasado. Pero tras un período inicial de soledad y depresión, Selkirk se adaptó a sus circunstancias, y se dedicó a cazar cabras, recolectar mariscos y frutas, recoger las verduras que habían plantado sus predecesores, hacer fuego frotando dos palos, fabricarse ropas con pieles de cabra y mantener a raya las ratas domesticando unas crías de gato para que compartieran sus habitaciones.
Selkirk fue rescatado por fin en 1709 por un barco británico que, curiosamente, era pilotado por el capitán del Cinque Ports. La tripulación quedó asombrada por aquel salvaje vestido con pieles que llevaba tanto tiempo solo que su inglés era casi ininteligible. Tras ayudar a aprovisionar el barco de fruta y carne de cabra, Selkirk embarcó de vuelta a Inglaterra. Allí se alió con un escritor para producir un popular relato de sus aventuras, The Englishman, que supuestamente inspiró el Robinson Crusoe de Daniel Defoe.[22] Sin embargo, Selkirk nunca logró adaptarse a la vida sedentaria en tierra firme. Volvió a navegar en 1720, y murió de unas fiebres un año más tarde en la costa africana.
Fueron las contingencias del tiempo y del carácter las que produjeron la historia de Selkirk. Pero la contingencia es también la lección de otra gran historia, la de los habitantes no humanos de las islas de Juan Fernández y de otras islas como ésta. Pues aunque Selkirk no lo supiera, Más a Tierra (hoy isla de Alexander Selkirk) estaba habitada por los descendientes de antiguos náufragos: los Robinson Crusoes de plantas, aves e insectos que arribaron a la isla por accidente miles de años antes que Selkirk. Sin saberlo, vivía en un laboratorio de la evolución.
En la actualidad las tres islas de Juan Fernández son un museo viviente de plantas y animales exóticos y raros, con numerosas especies endémicas, es decir, que no se encuentran en ningún otro lugar del mundo. Entre ellas se cuentan cinco especies de aves (incluido un colibrí gigante de unos doce centímetros y color pardo rojizo, el espectacular picaflor de Juan Fernández, que se encuentra en estado crítico de conservación), 126 especies de plantas (incluidos muchos miembros peculiares de la familia de los girasoles), un lobo de mar y un puñado de insectos. Ninguna otra superficie comparable en todo el mundo contiene tantas especies endémicas. Pero la isla es igualmente notable por lo que le falta: no acoge ni una sola especie de anfibio, reptil o mamífero, unos grupos que son comunes en los continentes de todo el mundo. Esta pauta de formas de vida extrañas y eflorescentes, con la sorprendente ausencia de muchos grandes grupos, se repite una y otra vez en las islas oceánicas. Y, como veremos, aporta importantes indicios a favor de la evolución.
Fue Darwin quien primero examinó a fondo estas pautas. Gracias a sus propios viajes de juventud en el HMS Beagle y a su voluminosa correspondencia con científicos y naturalistas, comprendió que era necesario apelar a la evolución para explicar no sólo los orígenes y formas de las plantas y animales sino también su distribución por el planeta. Estas distribuciones planteaban numerosas preguntas. ¿Por qué las islas oceánicas tienen floras y faunas tan extrañas y desequilibradas en comparación con los grupos de especies que se encontraban en los continentes? ¿Por qué casi todos los mamíferos autóctonos de Australia eran marsupiales, mientras que los placentarios predominaban en el resto del mundo? Y si las especies habían sido creadas, ¿por qué el creador había colocado en regiones distantes con un terreno y un clima parecidos, como los desiertos de África y América, especies semejantes en su apariencia pero que mostraban otras diferencias más fundamentales?
Otros antes que Darwin habían cavilado sobre estas preguntas y habían sentado los cimientos para su propia síntesis intelectual, una síntesis que Darwin consideraba tan importante que le dedicó dos capítulos enteros en El origen. Estos capítulos suelen considerarse el documento fundacional del campo de la biogeografía, la disciplina que estudia la distribución de las especies en la Tierra. La explicación evolutiva de la geografía de la vida que propuso entonces resultó ser en gran medida correcta, y sólo ha sido refinada y apoyada por una legión de estudios posteriores. La evidencia biogeográfica a favor de la evolución es en la actualidad tan poderosa que jamás me he encontrado con un libro, artículo o conferencia creacionista que haya intentado refutarla. Los creacionistas simplemente fingen que esa evidencia no existe.
Irónicamente, las raíces de la biogeografía se hunden en la religión. Los primeros «teólogos naturales» intentaron reconciliar la distribución de los organismos con el relato del arca de Noé de la Biblia. Todos los animales vivos se consideraban descendientes de las parejas que Noé llevó a bordo, parejas que viajaron hasta los lugares que ocupan en la actualidad desde el lugar donde quedó varada el arca después del diluvio (que tradicionalmente se supone que fue cerca del monte Ararat, en el levante de Turquía). Pero esta explicación tiene problemas evidentes. ¿Cómo se las arreglaron los canguros y las lombrices de tierra gigantes para cruzar los océanos hasta llegar a su hogar actual en Australia? ¿No se habría apresurado la pareja de leones a comerse a los antílopes? A medida que los naturalistas fueron descubriendo nuevas especies de plantas y animales, hasta los más recalcitrantes creyentes se dieron cuenta de que ninguna barca podía contenerlas a todas, y eso sin contar con su alimento y agua para un viaje de seis semanas.
Surgió entonces otra teoría: la de las creaciones múltiples distribuidas por toda la superficie de la Tierra. A mediados del siglo XIX, el prestigioso zoólogo Louis Agassiz, entonces en la Universidad de Harvard, afirmó que «no sólo eran las especies inmutables y estáticas, sino que también lo eran sus distribuciones, permaneciendo cada una en el lugar de su creación o cerca de él». Pero varios avances, sobre todo el número cada vez mayor de fósiles que refutaban la idea de que las especies fuesen «inmutables y estáticas», hicieron que tampoco esta idea pudiera sostenerse. Geólogos como Charles Lyell, el amigo y mentor de Charles Darwin, comenzaron a hallar indicios de que la Tierra no sólo era muy vieja, sino que además era cambiante. Durante el viaje del Beagle, el propio Darwin descubrió fósiles de conchas de moluscos a gran altitud en los Andes, lo que demostraba que lo que hoy es una montaña, en otro tiempo había estado bajo el mar. Las tierras podían ascender o hundirse, y los continentes que vemos en la actualidad podían haber sido mayores o menores en el pasado. Y quedaban además todas aquellas preguntas sin respuesta sobre la distribución de las especies. ¿Por qué la flora del sur de África se parecía tanto a la del sur del continente americano? Algunos biólogos propusieron que todos los continentes habían estado conectados en otro tiempo a través de gigantescos istmos, pero no existía ningún indicio de que hubieran existido (Darwin se quejó a Lyell de que estos istmos se habían hecho aparecer «con la misma facilidad con la que un cocinero hace unas tortas»).
En un intento por sortear estas dificultades, Darwin propuso su propia teoría: que las distribuciones de las especies no se explican por la creación, sino por la evolución. Si las plantas y los animales dispusieran de medios para dispersarse a gran distancia y, después de dispersarse, pudieran evolucionar, entonces, unido esto a algunos cambios antiguos en la Tierra, como los períodos de expansión glacial, podrían explicarse muchas de las peculiaridades de la biogeografía que habían desconcertado a sus predecesores.
Al final, Darwin resultó tener razón, aunque no en todo. Es cierto que muchas de las observaciones de la biogeografía cobran sentido si suponemos dispersión, evolución y una Tierra cambiante. Pero no todas. Las grandes aves no voladoras, como los avestruces, los ñandúes y los emúes, viven en África, América del Sur y Australia, respectivamente. Si todos hubieran tenido un mismo antepasado común, ¿cómo podía haberse dispersado tan ampliamente? ¿Y por qué el este de China y el este de América del Norte, dos regiones muy alejadas, comparten plantas, como los tuliperos y el dragón fétido, que no aparecen en las tierras que se extienden entre ellas?
Hoy disponemos de muchas de las respuestas que escaparon a Darwin gracias a dos avances que él no podía siquiera imaginar: la teoría de la deriva continental y la taxonomía molecular. Darwin comprendió que la Tierra había cambiado con el tiempo, pero no tenía ni idea de cuánto había cambiado realmente. Desde la década de 1960, los científicos saben que la geografía antigua del mundo era muy distinta de la actual, que unos ingentes supercontinentes se han desplazado, unido y separado después en fragmentos.[23]
Además, desde hace unos cuarenta años, hemos ido acumulando información de secuencias de ADN y proteínas que no sólo nos dicen cuáles son las relaciones evolutivas entre las especies, sino también el momento aproximado en que divergieron de sus antepasados comunes. La teoría de la evolución predice, y los datos lo apoyan, que a medida que las especies divergen de sus antepasados comunes, sus secuencias de ADN cambian de manera más o menos lineal con el tiempo. Podemos utilizar este «reloj molecular», calibrado con antepasados fósiles de especies existentes, para estimar el momento de divergencia de especies que tienen registros fósiles pobres.
Con la ayuda del reloj molecular podemos establecer las correspondencias entre las relaciones evolutivas entre especies y los movimientos conocidos de los continentes, además de los movimientos de los glaciares y la formación de auténticos puentes de tierra como el istmo de Panamá. Esto nos dice si los orígenes de las especies concurren en el tiempo con el origen de nuevos continentes y hábitats. Estas innovaciones han transformado la biogeografía en una gran historia de detectives: usando una serie de instrumentos y con un conjunto de hechos aparentemente inconexos, los biólogos deducen por qué las especies viven donde lo hacen. Hoy sabemos, por ejemplo, que las similitudes entre las plantas africanas y suramericanas no son sorprendentes, pues sus antepasados habitaron en otro tiempo en un supercontinente, Gondwana, que hace unos 170 millones de años se comenzó a separar en varios trozos (los actuales África, América del Sur, India, Madagascar y Antártida).
Cada uno de los pequeños trabajos de detective ya resueltos apoya el hecho de la evolución. Si las especies no hubieran evolucionado, sus distribuciones geográficas, tanto las actuales como las de sus fósiles, carecerían de sentido. Examinaremos primero las especies que viven en los continentes y luego las de las islas, pues estas dos geografías tan distintas aportan distintos indicios de la evolución.
Continentes
Comencemos por una observación que habrá llamado la atención de cualquier persona que haya viajado mucho. Cuando se visitan dos áreas con un terreno y un clima parecidos, se encuentran diferentes tipos de organismos. Tomemos el caso de los desiertos. Muchas plantas del desierto son suculentas: presentan una combinación de caracteres adaptativos como los tallos gruesos y carnosos para almacenar agua, las espinas para disuadir a los herbívoros y las hojas reducidas o ausentes para reducir la pérdida de agua. Pero cada desierto tiene un tipo de suculentas distinto. En el continente americano, las suculentas pertenecen a la familia de los cactus. Pero los desiertos de Asia, Australia y África no tienen cactus autóctonos, y en ellos las suculentas pertenecen a una familia distinta, la de las lechetreznas o euforbias. Pueden distinguirse los dos tipos de suculentas por las flores y por la savia, que es clara y acuosa en los cactus, y lechosa y amarga en las lechetreznas. Sin embargo, pese a estas diferencias fundamentales, cactus y euforbias pueden tener una apariencia muy semejante. Tengo plantas de los dos tipos en las macetas del alféizar, y mis visitas son incapaces de distinguirlas sin leer las etiquetas.
¿Por qué un creador habría de colocar plantas que son fundamentalmente distintas pero tienen aspecto muy parecido en áreas distantes del mundo que parecen ser ecológicamente idénticas? ¿No tendría más sentido poner las mismas especies de plantas en las regiones que tienen el mismo tipo de suelo y el mismo clima?
Podría replicarse que, si bien los desiertos parecen idénticos, son hábitats que difieren en aspectos sutiles pero importantes, y que los cactus y las euforbias se crearon de manera que se adecuaran a la perfección a sus hábitats respectivos. Pero esta explicación no vale, porque cuando se introducen cactus en los desiertos del Viejo Mundo, donde no se encuentran de manera natural, no tienen ningún problema para crecer. La chumbera, por ejemplo, que es un cactus originario de América del Norte, se introdujo en Australia a principios del siglo XIX con la intención de que los colonos pudieran extraer un tinte rojo de las cochinillas que se alimentan de la planta (éste es el tinte que da a las alfombras persas su intenso color rojo carmesí). A la entrada del siglo XX la chumbera se había extendido con tanta rapidez que ya era una grave plaga que había malogrado miles de hectáreas de tierras agrícolas y estimulado programas de erradicación tan drásticos como ineficaces. La planta no pudo controlarse hasta 1926 con la introducción de la polilla del nopal (Cactoblastis cactorum), cuyas orugas devoran el cactus, en lo que constituye el primer ejemplo de control biológico, y uno de los de mayor éxito. Está claro, pues, que las chumberas, que son cactus, pueden crecer estupendamente en los desiertos australianos aunque en éstos las suculentas autóctonas sean euforbias.
El ejemplo más famoso de especies distintas que desempeñan papeles parecidos es el de los mamíferos marsupiales, que en la actualidad se encuentran sobre todo en Australia (la zarigüeya es una excepción), y los mamíferos placentarios, que predominan en el resto del mundo. Los dos grupos presentan importantes diferencias anatómicas, sobre todo en su sistema reproductor (casi todos los marsupiales tienen bolsa, el marsupio, y paren crías muy poco desarrolladas, mientras que los placentarios poseen placentas que permiten que las crías nazcan con un grado de desarrollo mucho más avanzado). No obstante, en otros aspectos algunos marsupiales y placentarios muestran un parecido sorprendente. Hay topos marsupiales de aspecto y comportamiento casi igual al de los topos placentarios, ratones marsupiales que se parecen a los ratones placentarios, está el falangérido de azúcar, un marsupial que planea de árbol en árbol del mismo modo que las ardillas voladoras, y hay hormigueros marsupiales que hacen exactamente lo mismo que los osos hormigueros suramericanos (Figura 20).
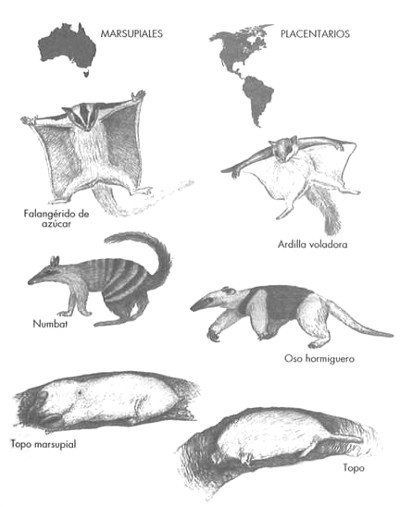
FIGURA 20. Evolución convergente en los mamíferos. Los hormigueros, falangéridos y topos marsupiales evolucionaron en Australia de manera independiente de sus equivalente placentarios de América; sus formas, sin embargo, son notablemente parecidas.
Una vez más, hay que preguntarse: si los animales hubieran sido creados especialmente, ¿por qué habría de producir el creador en continentes distintos unos animales fundamentalmente distintos que sin embargo tienen aspecto y comportamiento tan parecidos? No es que los marsupiales sean inherentemente superiores a los placentarios en Australia, porque a los mamíferos placentarios introducidos en este continente les ha ido muy bien. Los conejos, sin ir más lejos, se han convertido en una plaga tan seria en Australia que están desplazando a marsupiales autóctonos como el bilbi (un pequeño mamífero con unas orejas marcadamente largas). Para ayudar a recoger fondos para la erradicación del conejo, los conservacionistas están haciendo campaña para que se abandone el conejo de Pascua a favor del bilbi de Pascua, así que cada primavera los bilbis de chocolate llenan los estantes de los supermercados australianos.
Ningún creacionista, sea del ramo del arca de Noé o de cualquier otro, ha ofrecido jamás una explicación razonable de por qué algunos tipos de animales distintos tienen formas tan parecidas en lugares distintos. No les queda otra que apelar a los inescrutables caprichos del creador. La evolución, en cambio, explica estas observaciones invocando un proceso bien descrito conocido como evolución convergente. Es muy sencillo. Las especies que habitan en hábitats parecidos están sometidos a presiones de selección parecidas de su entorno, así que su evolución los lleva a desarrollar adaptaciones parecidas, es decir a converger, y acaban pareciéndose y comportándose de manera muy similar aunque no están emparentadas. Pero estas especies conservan diferencias esenciales que delatan su lejana ascendencia. (Un ejemplo famoso de convergencia es la coloración blanca de camuflaje que comparten diversos animales árticos como el oso polar y el búho nival.) El antepasado de los marsupiales colonizó Australia, mientras que los placentarios dominaron el resto del mundo. Tanto unos como otros se escindieron en multitud de especies, adaptadas a hábitats distintos. Si uno sobrevive y se reproduce mejor porque excava túneles bajo el suelo, la selección natural le reducirá los ojos y le proveerá de largas garras excavadoras, sin importar que se trate de un placentario o de un marsupial. Pero cada uno retendrá algunos rasgos característicos de sus antepasados.
Los cactus y las euforbias también muestran caracteres convergentes. El antepasado de las euforbias colonizó el Viejo Mundo, y el de los cactus el continente americano. Las especies que acabaron viviendo en los desiertos evolucionaron hacia adaptaciones parecidas: si uno es una planta en un clima seco, lo mejor es ser duro, no tener hojas y tener un tallo grueso para almacenar agua. Así que la selección natural moldeó las euforbias y los cactus de forma parecida.
La evolución convergente demuestra cómo funcionan de manera conjunta tres aspectos de la teoría de la evolución: la ascendencia común, la especiación y la selección natural. La ascendencia común explica por qué los marsupiales australianos comparten determinados caracteres (por ejemplo, las hembras tienen dos vaginas y dos úteros), mientras que los mamíferos placentarios comparten un conjunto distinto de caracteres (por ejemplo, una placenta persistente). La especiación es el proceso por el que el antepasado común da origen a muchas especies descendientes distintas. Y la selección natural hace que cada especie esté bien adaptada a su medio. Si ponemos todo esto junto y añadimos el hecho de que áreas distantes del mundo pueden tener hábitats parecidos, obtenemos la evolución convergente y, por añadidura, una explicación sencilla de las principales pautas biogeográficas.
Por lo que respecta a cómo llegaron los marsupiales a Australia, eso es parte de otra historia evolutiva, una historia que, además, conduce al enunciado de predicciones contrastables. Los fósiles de marsupiales más antiguos no se han encontrado en Australia sino en América del Norte. A medida que los marsupiales evolucionaron, se fueron extendiendo hacia el sur hasta alcanzar hace unos 40 millones de años lo que hoy es la punta más meridional de América del Sur. Los marsupiales llegaron a Australia aproximadamente unos 10 millones de años más tarde, y comenzaron a diversificarse en las alrededor de doscientas especies que viven allí en la actualidad.
Pero ¿cómo pudieron cruzar el Atlántico meridional? La respuesta es que todavía no existía. En el momento de la invasión de los marsupiales, América del Sur y Australia estaban unidas al sur del supercontinente de Gondwana. Esta masa continental ya había comenzado a separarse, desabrochándose para formar el océano Atlántico, pero la punta de América del Sur todavía estaba conectada con lo que hoy es la Antártida, que a su vez estaba conectada con lo que hoy es Australia (véase la Figura 21). Como los marsupiales tuvieron que viajar por la tierra firme desde América del Sur hasta Australia, a la fuerza tuvieron que cruzar la Antártida. Así que podemos predecir que debería haber fósiles de marsupiales en la Antártida que daten de hace 30 a 40 millones de años.
Esta hipótesis era lo bastante sólida como para empujar a algunos científicos hasta la Antártida en busca de marsupiales fósiles. Y, efectivamente, los encontraron: hallaron más de una docena de especies de marsupiales (que pueden identificarse como tales por su característica dentición y sus mandíbulas) en la isla Seymour, cerca de la península Antártica. Esta área se encuentra justo en el antiguo paso libre de hielo entre América del Sur y la Antártida. Y los fósiles tienen la edad predicha: de 35 a 40 millones de años. Tras su descubrimiento en 1982, el paleontólogo polar William Zinsmeister clamó exultante: «Durante años y años hemos pensado que aquí tenía que haber marsupiales. Esto encaja con todas las suposiciones que se habían hecho sobre la Antártida. Hemos encontrado lo que esperábamos encontrar».
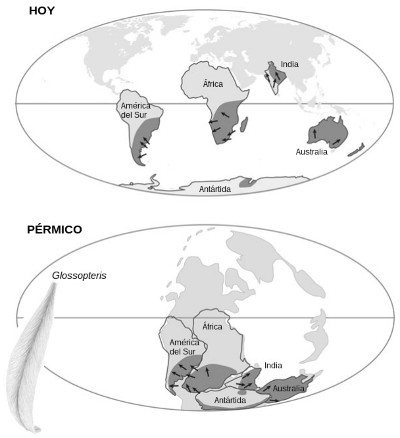
FIGURA 21. La deriva de los continentes explica la biogeografía evolutiva del género de árboles extintos Glossopteris. Arriba: la distribución actual de fósiles de Glossopteris (sombreado) está dividida en fragmentos distribuidos por varios continentes, lo que la hace difícil de interpretar. Las estrías que dejaron los glaciares en las rocas también resultan enigmáticas (flechas). Abajo: la distribución de Glossopteris cobra sentido durante el período Pérmico, cuando las tierras emergidas estaban unidas formando un supercontinente: las especies rodeaban el polo sur en una franja de clima templado. También las estrías glaciales pueden interpretarse ahora, pues todas apuntan hacia fuera del polo sur del Pérmico.
¿Y qué decir de los muchos casos de especies parecidas (pero no idénticas) que viven en hábitats parecidos pero en continentes distintos? El ciervo común vive en el norte de Europa, mientras que el uapití, que se le parece mucho, vive en América del Norte. Las ranas acuáticas sin lengua de la familia Pipidae viven en dos lugares muy distantes: el este de América del Sur y el África subtropical. Y ya hemos comentado el caso de las semejanzas en la flora del este de Asia y el este de América del Norte. Estas observaciones serian desconcertantes para los evolucionistas si los continentes siempre hubieran estado en los lugares que ocupan en la actualidad. Hubiera sido imposible entonces la dispersión de una magnolia de China a Alabama, de unas ranas de agua dulce entre África y América del Sur o que el ciervo ancestral hubiera pasado de Europa a América del Norte. Pero hoy sabemos perfectamente cómo se produjo esta dispersión: gracias a la existencia de antiguas conexiones de tierra firme entre los continentes. (Muy distintas de los enormes puentes de tierra que habían imaginado los primeros biogeógrafos.) Asia y América del Norte estuvieron en otro tiempo conectados por el puente de Beringia, a través del cual colonizaron este continente plantas y animales, incluidos los humanos. Y América del Sur y África fueron en otro tiempo parte de Gondwana.
Cuando los organismos se dispersan y colonizan con éxito una nueva área, con frecuencia evolucionan. Y esto nos lleva a otra predicción que ya enunciamos en el capítulo 1. Si se ha producido evolución, las especies que viven en un área determinada deberían descender de especies anteriores que habían habitado en el mismo lugar. Así que si excavamos en lechos poco profundos de las rocas de ese lugar, deberíamos encontrar fósiles que se asemejen a los organismos que pisan ese suelo en la actualidad.
Y así es. ¿Dónde encontramos los fósiles de canguro que más se parecen a los actuales? En Australia. Está también el caso de los armadillos del Nuevo Mundo. Los armadillos son unos mamíferos únicos por llevar por armadura (de ahí el nombre) un caparazón duro. Viven en todo el continente americano. ¿Dónde encontramos los fósiles que más se les parecen? En América, que fue el hogar de los gliptodontes, unos mamíferos herbívoros con caparazón y aspecto de armadillo recrecido. Algunos alcanzaban el tamaño de un escarabajo de Volkswagen, con una tonelada de peso, una armadura de unos cinco centímetros de grosor y unas bolas espinosas en la cola que blandían como mazas. El creacionismo lo tiene difícil para explicar todas estas observaciones: para hacerlo, tendría que proponer que en todo el mundo se ha ido produciendo un número inacabable de extinciones y creaciones sucesivas, y que en cada conjunto de nuevas creaciones, las especies se parecían a las que habían vivido anteriormente en el mismo lugar. Eso queda muy lejos del arca de Noé.
Que los ancestros fósiles y sus descendientes aparezcan en los mismos lugares conduce a una de las predicciones más famosas de la historia de la biología evolutiva: la hipótesis planteada por Darwin en El origen del hombre (1871) de que los humanos evolucionaron en África:
Es natural preguntarse entonces cuál fue el lugar de nacimiento del hombre en aquel estadio de la descendencia en el que nuestros progenitores divergieron de los catarrinos [los monos y simios del Viejo Mundo]. El hecho de que pertenecieran a este grupo muestra con claridad que habitaban en el Viejo Mundo; pero no en Australia ni en ninguna isla oceánica, como podemos inferir de las leyes de la distribución geográfica. En cada gran región del mundo los mamíferos existentes están estrechamente emparentados con las especies extintas de la misma región. Por consiguiente, es probable que África estuviera habitada con anterioridad por simios hoy extintos afines al gorila y al chimpancé; y como estas dos especies son en la actualidad las más parecidas al hombre, es algo más probable que nuestros primeros progenitores vivieran en el continente africano que en ningún otro lugar.
Cuando Darwin hizo esta predicción, nadie había visto todavía ningún fósil de los primeros humanos. Como veremos en el capítulo 8, los primeros restos no se descubrieron hasta 1924 y, como era de esperar, se hallaron en África. La profusión de fósiles de transición entre los simios y los humanos descubiertos desde entonces, de los que los más antiguos han aparecido siempre en África, no deja dudas de que Darwin tenía razón.
La biogeografía no se limita a hacer predicciones; también ha resuelto algunos enigmas. He aquí uno relativo a los glaciares y los árboles. Los geólogos saben desde hace tiempo que todos los continentes y subcontinentes meridionales experimentaron un intenso período de glaciaciones durante el período Pérmico, hace unos 290 millones de años. Sabemos que es así porque, a medida que se desplazan los glaciares, las rocas y cantos que arrastran dejan unas estrías características causadas por la fricción contra la roca madre. La dirección de estas estrías nos dice hacia dónde se movían los glaciares.
El examen de las rocas del Pérmico en las tierras meridionales revela unas estrías muy extrañas. Los glaciares parecen haberse originado en áreas como África central, que en la actualidad son muy cálidas y, lo que resulta todavía más confuso, parecen haberse desplazado del mar al continente. (Véase la dirección de las flechas en la Figura 21.) Todo esto parece imposible: los glaciares sólo se forman en climas constantemente fríos y sobre la tierra firme, donde las continuas nevadas se van compactando y formando hielo que comienza a moverse bajo su propio peso. ¿Cómo se explican entonces los dibujos de aspecto caprichoso que forman las estrías de los glaciares y su aparente origen en el mar?
Hay otra pieza más en este rompecabezas, pero no relacionada con la distribución de las estrías sino de unas especies de árboles, las del género Glossopteris. Estas coníferas tenían hojas en forma de lengua en lugar de agujas (glossa significa «lengua» en griego). Glossopteris fue una de las plantas que dominaron la flora del Pérmico. Por varias razones, los botánicos creen que eran árboles de hoja caduca, que dejaban caer cada otoño para producirlas de nuevo a la primavera siguiente, pues tenían anillos de crecimiento y características especializadas que indican que las hojas estaban programadas para separarse del árbol. Estos y otros rasgos sugieren que Glossopteris vivía en áreas templadas con inviernos fríos.
Cuando se sitúa sobre un mapa la distribución de los fósiles de Glossopteris en el hemisferio sur, que es la única región donde se han hallado (Figura 21), se observa un dibujo extraño formado por manchas inconexas en varios continentes. Esta distribución no puede explicarse por dispersión, pues las semillas de Glossopteris eran grandes y pesadas, y casi con toda seguridad no flotaban. ¿Podríamos tener aquí un caso de creación de la planta en distintos continentes? No tan rápido.
Los dos enigmas se resuelven cuando se atiende a la posición que los continentes actuales ocupaban a finales del Pérmico (Figura 21), cuando estaban unidos como piezas de un puzle formando un único continente: Gondwana. Cuando se unen las piezas, la posición de las estrías de los glaciares y la distribución de los árboles cobran sentido de inmediato. Ahora las estrías apuntan hacia afuera del centro de la Antártida, que resulta ser la parte de Gondwana que pasó por el polo sur durante el Pérmico. La nieve debió producir entonces extensos glaciares que se desplazaban alejándose de esta área y produciendo, por tanto, estrías en las direcciones observadas. Y cuando en el mapa de Gondwana se superpone la distribución de Glossopteris, el dibujo que aparece deja de ser caótico: ahora las manchas están conectadas formando un anillo alrededor de los márgenes de los glaciares. Éstas son precisamente las áreas de clima fresco donde cabe encontrar árboles de hoja caduca.
Así que no fueron los árboles los que migraron de un continente a otro, cruzando grandes distancias, sino que fueron los continentes mismos los que se movieron, llevando los árboles con ellos. Estos rompecabezas cobran sentido a la luz de la evolución, mientras que el creacionismo carece de explicaciones para el dibujo de las estrías glaciales o la peculiar distribución disyunta de Glossopteris.
Esta historia tiene una nota triste. Cuando en 1912 se encontró el grupo de exploradores dirigidos por Robert Scott, que habían muerto congelados después de un intento fallido de convertirse en los primeros en alcanzar el polo sur (el noruego Roald Amundsen llegó un poco antes), junto a sus cuerpos se encontraron unos quince kilos de fósiles de Glossopteris. Pese a haber abandonado buena parte de su equipaje en un intento desesperado por mantenerse con vida, el grupo había seguido arrastrando estas pesadas rocas en trineos de mano, conocedores sin duda de su valor científico. Eran los primeros especímenes de Glossopteris descubiertos en la Antártida.
Las pruebas de la evolución que aporta la distribución de la vida sobre los continentes es sólida, pero más aún lo es, como veremos enseguida, la que aporta la distribución de la vida en las islas.
Islas
Darse cuenta de que la distribución de las especies en las islas proporciona una prueba concluyente de la evolución fue una de las mayores hazañas detectivescas de la historia de la biología. También ésta se la debemos a Darwin, cuyas ideas todavía pesan mucho en el campo de la biogeografía. En el capítulo 12 de El origen, Darwin presenta un hecho tras otro recogidos meticulosamente durante años de observaciones y correspondencia, hasta construir su caso como un brillante abogado. Cuando enseño a mis alumnos las pruebas de la evolución, ésta es mi clase preferida, una historia de misterio que parte de la acumulación de datos dispares que al cabo de la hora se resuelve en un sólido argumento en favor de la evolución.
Pero antes de presentar la evidencia, necesitamos distinguir entre dos tipos de islas. El primero es el de las islas continentales, aquellas que en otro tiempo estuvieron conectadas al continente pero quedaron separadas de él por una subida del nivel del mar o por el movimiento de las placas continentales. A este tipo pertenecen, entre muchas otras, las islas Británicas, Japón, Sri Lanka, Tasmania y Madagascar. Algunas son muy antiguas (Madagascar se apartó de África hace unos 160 millones de años), otras mucho más jóvenes (Gran Bretaña se separó de Europa hace unos 300.000 años, probablemente a causa de la rotura catastrófica de la presa natural que contenía un gran lago situado al norte). Las islas oceánicas, por otro lado, son aquellas que nunca estuvieron conectadas con un continente; se alzaron desde el fondo del mar, al principio totalmente desprovistas de vida, por la erupción de volcanes o el crecimiento de arrecifes coralinos. Se incluye en este grupo a las islas de Hawái, el archipiélago de Galápagos, Santa Elena y el grupo de Juan Fernández, descrito al principio de este capítulo.
El argumento a favor de la evolución que se fundamenta en las islas comienza con la siguiente observación: en las islas oceánicas faltan muchos tipos de especies autóctonas que encontramos tanto en los continentes como en las islas continentales. Tomemos el caso de Hawái, un archipiélago tropical cuyas islas ocupan unos 17.000 kilómetros cuadrados, algo menos de la superficie del estado de Massachusetts. Aunque estas islas están bien provistas de aves, plantas e insectos autóctonos, carecen por completo de peces de agua dulce, anfibios, reptiles y mamíferos terrestres que no sean introducidos. Santa Elena, la isla de Napoleón, y el archipiélago de Juan Fernández carecen de estos mismos grupos, y tienen en cambio abundantes plantas, aves e insectos. Las islas Galápagos tienen unos pocos reptiles autóctonos (iguanas terrestres y marinas, además de las famosas tortugas gigantes), pero carecen como las otras de mamíferos, anfibios o peces de agua dulce autóctonos. Una y otra vez, en las islas oceánicas que salpican el Pacífico, el Atlántico sur y el océano Índico, se sigue la misma pauta: faltan grupos. Más aún, faltan siempre los mismos grupos.
A primera vista, estas ausencias resultan extrañas. En la más pequeña parcela de territorio de un continente o isla continental de la región tropical, por ejemplo en Perú, Nueva Guinea o Japón, se encuentran indefectiblemente abundantes especies autóctonas de peces, anfibios, reptiles y mamíferos.
Tal como observó Darwin, esta disparidad es difícil de explicar bajo un prisma creacionista: «Quien admita la doctrina de la creación separada para cada especie, tendrá que admitir que para las islas oceánicas no fue creado un número suficiente de plantas y animales bien adaptados». Pero ¿cómo sabemos que los mamíferos, anfibios, peces de agua dulce y reptiles son realmente adecuados para las islas oceánicas? A lo mejor el creador no las puso porque en ellas no podrían desarrollarse. Una respuesta obvia es que las islas continentales sí tienen esos animales, y ¿por qué habría de poner el creador tipos distintos de animales en las islas dependiendo de si son oceánicas o continentales? Cómo se formó la isla no debería importar. Pero Darwin acaba la frase antes citada con una respuesta aún mejor: «… pues el hombre involuntariamente las ha poblado de modo mucho más completo y perfecto que lo hizo la naturaleza».
Dicho de otro modo, los mamíferos, los anfibios, los peces de agua dulce y los reptiles a menudo se las arreglan muy bien en las islas oceánicas donde el hombre las ha introducido. De hecho, es frecuente que dominen a las especies autóctonas, barriéndolas en algunos casos. Los cerdos y cabras introducidos en Hawái han hecho pasto de las plantas nativas. Las ratas y las mangostas introducidas han destruido o puesto en peligro de extinción a muchas de las espectaculares aves de Hawái. El sapo de la caña, un enorme anfibio venenoso nativo de la América tropical, fue introducido en Hawái en 1932 para controlar los escarabajos de la caña de azúcar. En la actualidad se han convertido en una plaga: se reproducen prolíficamente y matan a los gatos y perros que los toman por alimento. Las islas Galápagos no tienen anfibios autóctonos, pero la ranita arbórea ecuatoriana, introducida en 1998, se ha establecido en tres de las islas. En Sao Tomé, una isla volcánica situada frente a la costa occidental de África donde capturo moscas de la fruta para mi propia investigación, se han introducido cobras negras (quizá de manera accidental) desde el continente africano, y les va tan bien en la isla que nos hemos visto obligados a dejar de trabajar en determinadas áreas donde las cobras son tan numerosas que en un solo día nos podríamos encontrar con varias de estas serpientes mortales y agresivas. Los mamíferos terrestres también pueden vivir bien en las islas: las cabras introducidas en Más a Tierra ayudaron a Alexander Selkirk a sobrevivir, y también viven muy bien en Santa Elena. Dondequiera que se vaya, la historia es la misma: los humanos introducen especies en islas oceánicas donde antes no existían, y estas especies desplazan o destruyen las formas nativas. Ahí queda el argumento de que las islas oceánicas son de algún modo inadecuadas para mamíferos, anfibios, reptiles y peces.
El siguiente paso del argumento es el siguiente: aunque las islas oceánicas carecen de muchas clases básicas de animales, en las que se encuentran, lo hacen a menudo con una profusión de especies semejantes. Veamos de nuevo el caso de las Galápagos. En sus trece islas hay un total de veintiocho especies de aves que no se encuentran en ningún otro lugar. Y de estas veintiocho, catorce pertenecen a un mismo grupo de aves muy estrechamente emparentadas: los famosos pinzones de las Galápagos. Ningún continente o isla continental tiene una avifauna tan claramente dominada por pinzones. Pero pese a los caracteres que comparten como tales, los pinzones de las Galápagos son ecológicamente bastante diversos, con distintas especies especializadas en alimentos tan diferentes como insectos, semillas y huevos de otras especies. El «pinzón carpintero» es una de esas raras especies que utilizan herramientas, en su caso una espina de cactus o una ramita para sacar los insectos de los árboles. Los pinzones carpinteros desempeñan el papel ecológico de los pájaros carpinteros, de los que no hay ninguna especie en las islas Galápagos. Y hay incluso un «pinzón vampiro» que picotea los cuartos traseros de las aves marinas para luego lamer la sangre.
Hawái posee también su propia radiación de aves, más espectacular si cabe: la de los trepadores mieleros hawaianos. Cuando los polinesios llegaron a Hawái hace unos mil quinientos años, encontraron alrededor de 140 especies de aves autóctonas (lo sabemos gracias a los estudios de «subfósiles» de aves, es decir, de huesos preservados en antiguos vertederos de desechos y en tubos de lava). Unas sesenta de estas especies, casi la mitad de la avifauna del archipiélago, eran trepadores mieleros, y todos habían descendido de un única especie ancestral de pinzón que habían llegado a la isla hace unos 4 millones de años. Por desgracia, en la actualidad sólo quedan veinte especies de trepadores hawaianos, todas ellas amenazadas. El resto fue extinguido por la caza, la pérdida de hábitat y los depredadores introducidos por el hombre, como las ratas y las mangostas. Pero incluso los pocos trepadores que quedan exhiben una fabulosa diversidad de roles ecológicos, tal como muestra la Figura 22. El pico de un ave nos dice mucho sobre su dieta. Algunas especies tienen picos curvos para chupar el néctar de las flores, otras tienen picos robustos, como los de los loros, para partir semillas duras o estrujar tallos, otras tienen picos curvados para picotear los insectos entre el follaje, y aun otras tienen picos ganchudos para extraer insectos de los troncos de los árboles, en el papel que habitualmente desempeñan los pájaros carpinteros. Como en las Galápagos, vemos un grupo que se encuentra representado en exceso, con especies que ocupan los nichos que en los continentes y las islas continentales ocupan especies muy diferentes.
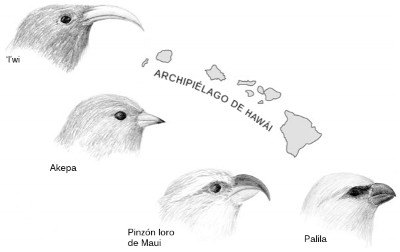
FIGURA 22. Una radiación adaptativa: algunas especies emparentadas de trepadores mieleros hawaianos que evolucionaron después de que su antepasado común, un ave con aspecto de pinzón, colonizara las islas. Cada pinzón tiene un pico que le permite explotar una fuente de alimento distinta. El fino pico del ‘i’wi lo ayuda a chupar el néctar de largas flores tubulares, el akepa tiene un pico ligeramente cruzado con el que puede abrir los brotes y los capullos en busca de insectos y arañas, el pinzón loro de Maui tiene un pico robusto para levantar la corteza de los árboles y partir los tallos y ramitas en busca de larvas de escarabajos, y el pico corto pero fuerte del palila le permite abrir vainas para extraer las semillas.
Las islas oceánicas también albergan radiaciones de plantas e insectos. Santa Elena carece de muchos grupos de insectos, pero da cobijo a docenas de especies de escarabajos pequeños y no voladores, sobre todo gorgojos de la madera. En Hawái, las moscas de la fruta del género Drosophila, que es el grupo que estudio, manifiestan una diversidad exuberante. Aunque las islas de Hawái representan tan sólo el 0,004 por 100 de la superficie de las tierras emergidas del planeta, contienen casi la mitad de las dos mil especies de Drosophila del mundo. También son ejemplos las notables radiaciones de plantas de la familia de los girasoles en el archipiélago de Juan Fernández y en Santa Elena, algunas de las cuales se han convertido en pequeños árboles leñosos. Sólo en las islas oceánicas pueden las pequeñas plantas con flor, libres de la competencia de arbustos y árboles mayores, evolucionar ellas mismas hasta convertirse en árboles.
Hasta aquí hemos pasado revista a dos conjuntos de hechos observados en las islas oceánicas: carecen de muchos grupos de especies que viven en los continentes y las islas continentales, y sin embargo los grupos que sí se encuentran en las islas oceánicas están repletos de especies similares. Juntas, estas dos observaciones muestran que, en comparación con otras áreas del mundo, la vida en las islas oceánicas está desequilibrada. Cualquier teoría biogeográfica que merezca ese nombre tiene que dar cuenta de este contraste.
Pero hay algo más. El siguiente cuadro presenta una lista de los principales grupos que suelen ser autóctonos de las islas oceánicas y de los que suelen faltar (Juan Fernández es uno de los grupos de islas que se ajustan a esta lista):
| Autóctonas | Faltantes |
|---|---|
|
Plantas Aves Insectos y otros artrópodos (por ejemplo, arañas) |
Mamíferos terrestres Reptiles Anfibios Peces de agua dulce |
¿Cuál es la diferencia entre las dos columnas? Basta con pensar un poco para ver la respuesta. Las especies de la primera columna pueden colonizar las islas oceánicas porque pueden dispersarse a grandes distancias; las especies de la segunda columna carecen de esa capacidad. Las aves pueden volar a gran distancia sobre el mar, llevando con ellas no sólo sus propios huevos, sino también las semillas de plantas que hayan comido (que pueden germinar en las propias deyecciones), parásitos en las plumas y pequeños organismos en el limo pegado a sus patas. Las plantas pueden llegar a las islas en forma de semillas, flotando por el mar a enormes distancias. Las semillas con pelos ganchudos o cubiertas pegajosas pueden viajar hasta las islas en las plumas de las aves. Las ligeras esporas de los helechos, los hongos y los musgos pueden desplazarse a mucha distancia arrastrados por el viento. Los insectos también pueden volar a las islas o dejarse arrastrar por el viento.
En cambio, los animales de la segunda columna tienen grandes dificultades para cruzar las grandes extensiones oceánicas. Los mamíferos terrestres y los reptiles son pesados y no pueden nadar muy lejos. Y la mayoría de los anfibios y peces de agua dulce sencillamente no pueden sobrevivir en el agua salada.
Así que los grupos de especies que hallamos en las islas oceánicas son precisamente aquellos que pueden llegar hasta ellas cruzando el mar desde tierras muy lejanas. Pero ¿qué pruebas tenemos de que lo hacen? Cualquier aficionado a las aves nos hablará de «visitantes» ocasionales encontrados a miles de kilómetros de su hábitat normal, víctimas de los vientos o de errores de orientación. Algunas aves han llegado incluso a establecer colonias en islas oceánicas en tiempos históricos. El calamoncillo americano, que durante mucho tiempo había sido un visitante accidental de la isla de Tristán da Cunha, en el Atlántico sur, por fin comenzó a anidar allí durante la década de 1950.
El propio Darwin realizó algunos experimentos sencillos pero elegantes para mostrar que las semillas de algunas especies de plantas conservaban la capacidad de germinar después de pasar mucho tiempo inmersas en agua del mar. Por otro lado, se han encontrado semillas de las Indias Occidentales en las lejanas costas de Escocia, adonde llegaron sin duda arrastradas por la corriente del golfo, y también se han encontrado «semillas de deriva» originarias de continentes o de otras islas en las costas de las islas del Pacífico sur. Las aves enjauladas pueden retener semillas de plantas en su sistema digestivo durante una semana o más, lo que demuestra la posibilidad de que las transporten a largas distancias. Y se ha conseguido muchas veces capturar insectos suspendidos en el aire utilizando trampas llevadas por aviones o barcos en lugares alejados de la tierra. Entre las especies recogidas se cuentan langostas, polillas, mariposas, moscas, pulgones y escarabajos. Ya en 1933, durante su viaje a través del Atlántico, Charles Lindbergh expuso al aire portaobjetos de microscopio en los que capturó numerosos microorganismos y partes de insectos. Muchas arañas se dispersan durante los estadios juveniles dejándose arrastrar por el aire gracias a unos «paracaídas» que elaboran con su seda; estos vagabundos se han encontrado a cientos de kilómetros de la tierra.
Los animales y las plantas también pueden viajar hasta las islas encima de «balsas» formadas por troncos y masas de vegetación que flotan a la deriva desde los continentes, por lo general desde las desembocaduras de los ríos. En 1995 una de estas grandes balsas, empujada quizá por un huracán, depositó su carga de quince iguanas en la isla caribeña de Anguilla, donde no habían existido hasta entonces, desde un lugar de origen situado a más de trescientos kilómetros de distancia. En Hawái se han encontrado troncos de abeto de Douglas, de América del Norte, y hasta Tasmania han llegado troncos originarios de América del Sur. Estas balsas explican la ocasional presencia de reptiles en islas oceánicas, como las iguanas y tortugas de las Galápagos.
Además, cuando se analiza qué tipo de insectos y plantas son autóctonos de las islas oceánicas, se ve que proceden de los grupos que son buenos colonizadores. La mayoría de los insectos son pequeños, pues son precisamente los que pueden ser llevados con facilidad por el viento. Comparados con las plantas herbáceas, los árboles son relativamente raros en las islas oceánicas, casi con certeza porque muchos árboles producen semillas pesadas que ni flotan ni son comidas por las aves. (Una excepción notable es el cocotero, de frutos grandes que flotan, una palmera que se encuentra en casi todas las islas de los océanos Pacífico e Índico.) De hecho, la rareza relativa de árboles explica por qué muchas plantas que en los continentes tienen porte herbáceo, en las islas han evolucionado hacia formas arbóreas.
Los mamíferos terrestres no son buenos colonizadores, y por eso faltan en las islas oceánicas. Pero no faltan todos los mamíferos. Esto nos lleva a comentar dos excepciones que confirman la regla. La primera fue observada ya por Darwin:
Aun cuando los mamíferos terrestres no existan en las islas oceánicas, los mamíferos aéreos existen en casi todas las islas. Nueva Zelanda posee dos murciélagos que no se encuentran en ninguna otra parte del mundo; la isla de Norfolk, el archipiélago de Viti, las islas Bonin, los archipiélagos de las Carolinas y de las Marianas, la isla de Mauricio poseen todas sus murciélagos peculiares. ¿Por qué la supuesta fuerza creadora —podría preguntarse— ha producido murciélagos y no otros mamíferos en las islas alejadas? Dentro de mi teoría esta pregunta puede contestarse fácilmente, pues ningún mamífero terrestre puede ser transportado a través de un gran espacio de mar; pero los murciélagos pueden volar y atravesarlo.
Y también hay mamíferos acuáticos en las islas. Hawái tiene uno, la foca monje de Hawái, que es endémica, y las islas de Juan Fernández tienen un lobo marino autóctono. Si los mamíferos autóctonos de las islas oceánicas no fueron creados, sino que descienden de ancestros colonizadores, cabe predecir que esos ancestros debían poder volar o nadar.
Es evidente que la dispersión a gran distancia de una especie determinada hasta una isla lejana no puede ser un evento frecuente. La probabilidad de que un insecto o un ave pueda no ya atravesar grandes extensiones de océano hasta aterrizar en una isla, sino establecer una población reproductora una vez allí (lo cual requiere que o bien llegue una hembra fecundada o al menos dos individuos de sexo opuesto), debe de ser muy reducida. Si la dispersión fuese más frecuente, la vida en las islas oceánicas se parecería bastante a la de los continentes y las islas continentales. No obstante, la mayoría de las islas oceánicas tienen edades que se miden en millones de años, tiempo suficiente para que se produzca alguna colonización. Como bien señalaba el zoólogo George Gaylord Simpson: «Todo evento que no sea absolutamente imposible… se hace probable si transcurre el tiempo suficiente». Veamos un caso hipotético. Supongamos que cierta especie sólo tiene una oportunidad en un millón de colonizar una isla en un año dado. Es fácil ver que después de un millón de años, hay una elevada probabilidad de que la isla haya sido colonizada al menos una vez: un 63 por 100, para ser precisos.[24]
Una última observación pone el broche a la cadena lógica que asegura el caso a favor de la evolución en las islas. Es ésta: con pocas excepciones, los animales y plantas de las islas oceánicas se parecen más que a otras a las especies que se encuentran en las tierras continentales más cercanas. Así pasa, por ejemplo, en las islas Galápagos, cuyas especies se parecen a las de la costa occidental de América del Sur. La similitud no puede explicarse con el argumento de que las islas y América del Sur tengan hábitats parecidos para unas especies creadas por un acto divino, porque las Galápagos son secas, volcánicas y sin árboles, muy distintas de los exuberantes trópicos que predominan en la costa americana. Darwin se muestra especialmente elocuente a este respecto:
El naturalista, al contemplar los habitantes de estas islas volcánicas del Pacífico, distantes del continente varios centenares de millas, tiene la sensación de que se encuentra en tierra americana. ¿Por qué ha de ser así? ¿Por qué las especies que se supone que han sido creadas en el archipiélago de los Galápagos y en ninguna otra parte han de llevar tan visible el sello de su afinidad con las creadas en América? Nada hay allí, ni en las condiciones de vida, ni en la naturaleza geológica de las islas, ni en su altitud o clima, ni en las proporciones en que están asociadas mutuamente las diferentes clases, que se asemeje mucho a las condiciones de la costa de América del Sur; en realidad, hay una diferencia considerable por todos estos conceptos… Hechos como éstos no admiten explicación de ninguna clase dentro de la opinión corriente de las creaciones independientes; mientras que, según la opinión que aquí se defiende, es evidente que las islas de los Galápagos estarían en buenas condiciones para recibir colonos de América, ya por medios ocasionales de transporte, ya —aun cuando yo no creo en esta teoría— por antigua unión con el continente … estos colonos estarían sujetos a modificación, delatando todavía el principio de la herencia de su primitivo lugar de origen.
Lo que es cierto de las Galápagos es cierto también de otras islas oceánicas. Los parientes más cercanos de las plantas y animales endémicos de Juan Fernández provienen de los bosques templados del sur del continente americano, el continente más cercano. La mayoría de las especies de Hawái son similares (no idénticas) a las de las regiones más cercanas, la indopacífica (Indonesia, Nueva Guinea, Fiji, Samoa y Tahití) y América. Ahora bien, si se tiene en cuenta los caprichos de los vientos y la dirección de las corrientes oceánicas, no cabe esperar que todos los colonizadores de las islas provengan de la fuente más cercana. El 4 por 100 de las especies vegetales de Hawái, por ejemplo, tienen sus parientes más cercanos en Siberia o en Alaska. Aun así, la similitud de las especies de las islas con las de los continentes más cercanos exige una explicación.
En resumen, las islas oceánicas poseen características que las distinguen de los continentes y de las islas continentales. Las islas oceánicas tienen biotas poco equilibradas, en las que faltan algunos de los grandes grupos de organismos, y además faltan los mismos grupos en las distintas islas. Pero los tipos de organismos que sí se encuentran en ellas a menudo comprenden muchas especies parecidas, una radiación, y son los tipos de especies, como las aves y los insectos, que pueden dispersarse más fácilmente atravesando grandes extensiones del océano. Además, las especies más parecidas a las que habitan en las islas oceánicas por lo general se encuentran en las tierras continentales más cercanas, aunque en ellas los hábitats sean distintos.
¿Cómo encajan todas estas observaciones? Cobran sentido a la luz de una simple explicación evolutiva: los habitantes de las islas oceánicas descienden de especies anteriores que colonizaron las islas, por lo general procedentes de continentes cercanos, en raros eventos de dispersión a larga distancia. Una vez allí, los colonos accidentales lograron formar muchas especies porque las islas oceánicas ofrecen muchos hábitats vacíos en los que faltan competidores y depredadores. Esto explica por qué la especiación y la selección natural se desboca en las islas, produciendo «radiaciones adaptativas» como la de los trepadores mieleros hawaianos. Todo encaja si se añade la dispersión ocasional, que sabemos que se produce, a los procesos darwinistas de selección, evolución, ascendencia común y especiación. En suma, las islas oceánicas demuestran todos los principios de la teoría de la evolución.
Es importante recordar que las islas continentales no suelen conformarse a estas pautas (veremos una excepción en un segundo), pues comparten especies con los continentes a los que en otro tiempo pertenecían. Las plantas y animales de Gran Bretaña, por ejemplo, forman un ecosistema mucho más equilibrado, con especies casi siempre idénticas a las de Europa continental. A diferencia de las islas oceánicas, las continentales se desgajaron con la mayoría de sus especies incorporadas.
Intentemos pensar ahora en una teoría que explique las pautas que hemos discutido apelando a actos especiales de creación de las especies de las islas oceánicas y continentales. ¿Por qué un creador habría de dejar a los anfibios, los mamíferos, los peces y los reptiles fuera de las islas oceánicas pero no de las continentales? ¿Por qué produjo un creador radiaciones de especies similares en las islas oceánicas, pero no en las continentales? ¿Y por qué las especies de las islas continentales se crearon para que se parecieran a las del continente más cercano?
No hay buenas respuestas a no ser que se suponga que el propósito del creador era hacer que pareciera que las especies habían evolucionado en islas. Nadie está dispuesto a abrazar esta respuesta, lo que explica que los creacionistas simplemente rehuyan la biogeografía de las islas.
Podemos enunciar ahora una última predicción. Las islas continentales muy antiguas, las que se hayan separado del continente hace millones de años, deberían presentar pautas evolutivas intermedias entre las de las islas continentales jóvenes y las islas oceánicas. Las islas continentales antiguas, como Madagascar o Nueva Zelanda, que se separaron de sus continentes hace 160 millones y 85 millones de años, respectivamente, quedaron aisladas antes de que hubieran evolucionado muchos grupos de primates y plantas modernas. Una vez estas islas se apartaron del continente, algunos de sus nichos ecológicos permanecieron desocupados. Esto abrió las puertas a que especies que evolucionaron más tarde lograsen colonizarlas con éxito y establecerse. Podemos predecir, por consiguiente, que estas viejas islas continentales deberían contener una flora y una fauna algo desequilibrada, con algunas de las peculiaridades biogeográficas de las islas oceánicas verdaderas.
Y, en efecto, eso es lo que encontramos. Madagascar es famosa por sus inusuales flora y fauna, que incluyen muchas plantas autóctonas y, por supuesto, sus lémures, los primates más primitivos, que sólo viven allí y cuyos ancestros, después de llegar a Madagascar hace unos 60 millones de años, experimentaron una radiación que dio origen a más de setenta y cinco especies endémicas. Nueva Zelanda también tiene muchas especies propias, de las que son especialmente célebres las aves no voladoras: el moa gigante, un monstruo de cuatro metros de altura que fue cazado hasta su extinción hacia 1500; el kiwi, y una especie de loro grande y no volador, el kakapo. Nueva Zelanda también muestra algo del «desequilibrio» de las islas oceánicas: sólo tiene unos pocos reptiles endémicos, sólo una especie de anfibio y dos mamíferos autóctonos, ambos murciélagos (aunque recientemente se ha hallado un pequeño mamífero fósil). También allí se produjo una radiación: hubo once especies de moa, todas extintas en la actualidad. Y, al igual que las islas oceánicas, las especies de Madagascar y Nueva Zelanda están emparentadas con las que se encuentran en los continentes más cercanos: África y Australia, respectivamente.
Epílogo
La principal lección de la biogeografía es que sólo la evolución puede explicar la diversidad de la vida en los continentes y las islas. Pero hay otra lección: la distribución de la vida en la Tierra refleja una mezcla de azar y necesidad. Azar, porque la dispersión de animales y plantas depende de los impredecibles caprichos de los vientos, las corrientes y la oportunidad de colonizar. Si a las Galápagos o a Hawái no hubieran llegado los primeros pinzones, quizá hoy veríamos en estas islas aves muy distintas. Si a Madagascar no hubiera llegado un animal parecido a los lémures, la isla (y probablemente la Tierra) no tendría hoy lémures. El tiempo y el azar bastan para explicar quién acaba náufrago en una isla, en lo que podríamos llamar «efecto Robinson Crusoe». Pero también está la necesidad. La teoría de la evolución predice que muchas de las plantas y animales que arriban a unos hábitats nuevos y desocupados evolucionarán y crecerán en ellos, formando nuevas especies que ocuparán nichos ecológicos. Y que normalmente tendrán sus parientes más cercanos en la isla o continente más cercano. Esto es lo que vemos una y otra vez. No puede entenderse la evolución si no se comprende su interacción única entre el azar y la necesidad, entre la oportunidad y la ley, una interacción que, como veremos en el siguiente capítulo, reviste una importancia crucial para entender la idea de la selección natural.
Pero las lecciones de la biogeografía van más allá, adentrándose en el dominio de la conservación biológica. Las plantas y animales de las islas se han adaptado a sus entornos aislados de otras especies que se encuentran en todos lados, sus competidores, depredadores y parásitos potenciales. Como las especies de las islas no experimentan la diversidad de la vida que se encuentra en los continentes, interaccionar con otros es algo que no se les da muy bien. Por eso los ecosistemas de las islas son frágiles, asolados con facilidad por especies foráneas que pueden destruir hábitats y acabar con especies. La peor es la humana, que no sólo tala los árboles y caza, sino que además trae consigo todo un séquito de destructivas chumberas, ovejas, cabras, ratas y sapos. Muchas de las especies únicas de las islas oceánicas ya se han extinguido, víctimas de las actividades humanas, y lamentablemente podemos decir con seguridad que muchas más desaparecerán en un futuro cercano. Durante nuestra vida quizá veamos al último de los trepadores mieleros de Hawái, o la extinción de los kakapos y kiwis de Nueva Zelanda, la reducción catastrófica de los lémures y la pérdida de muchas especies raras de plantas, quizá menos carismáticas, pero no por ello menos interesantes. Cada especie representa millones de años de evolución y, una vez desaparecida, nunca más podrá volver. Y cada una es un libro que contiene una historia única sobre el pasado. Perder cualquiera de ellas es perder una parte de la historia de la vida.