Capítulo 2
La corteza terrestre es un inmenso museo; pero las colecciones naturales han sido hechas a intervalos de tiempo inmensamente dilatados.
CHARLES DARWIN, El origen de las especies
La historia de la vida en la Tierra está escrita en las rocas. Se trata, qué duda cabe, de un libro de historia roto y desvencijado, del que sólo hallamos restos dispersos de lo que fueron sus hojas. Pero ahí está, y hay partes significativas que todavía son legibles. Los paleontólogos han trabajado sin cesar para reconstruir los indicios históricos tangibles de la evolución: el registro fósil.
Cuando admiramos alguno de los fósiles más impresionantes, como los grandes esqueletos de dinosaurios que adornan nuestros museos de historia natural, es fácil olvidar el enorme esfuerzo que ha representado descubrirlos, excavarlos, prepararlos y describirlos. Con frecuencia han sido necesarias largas, costosas y arriesgadas expediciones a los rincones más remotos e inhóspitos del planeta. Sin ir más lejos, uno de mis colegas de la Universidad de Chicago, Paul Sereno, estudia dinosaurios africanos, y muchos de los fósiles más interesantes que ha descubierto los ha encontrado justo en medio del desierto del Sahara. Él y sus compañeros de trabajo han tenido que habérselas con conflictos políticos, bandidos, enfermedades y, por descontado, con los rigores del propio desierto para descubrir nuevas e interesantes especies como Afrovenator abakensis y Jobaria tiguidensis, especímenes que han ayudado a reescribir la historia de la evolución de los dinosaurios.
Descubrimientos como éstos requieren una genuina dedicación a la ciencia, muchos años de trabajo meticuloso, persistencia y valentía, además de una saludable dosis de buena suerte. Pero muchos paleontólogos están dispuestos a arriesgar sus vidas por realizar descubrimientos como éstos. Para los biólogos, los fósiles son tan valiosos como las pepitas de oro. Sin ellos, no tendríamos más que un esbozo imperfecto de la evolución. Nos veríamos limitados a estudiar las especies vivas e intentar inferir sus relaciones evolutivas a partir de su parecido de forma, desarrollo y secuencia de ADN. Sabríamos, por ejemplo, que los mamíferos están más emparentados con los reptiles que con los anfibios, pero no sabríamos nada sobre cómo eran sus antepasados comunes. No tendríamos el menor indicio de la existencia de los grandes dinosaurios, algunos tan grandes como camiones, o de nuestros antepasados australopitecinos más antiguos, con un volumen craneano pequeño pero que ya caminaban erectos. Mucho de lo que nos gustaría saber sobre la evolución quedaría envuelto en el misterio. Por suerte, los avances de la física, la geología y la bioquímica, junto con la audacia y la perseverancia de científicos de todo el mundo, nos permiten disfrutar de esta valiosa mirada al pasado.
Cómo se hace el registro
Los fósiles se conocen desde los tiempos más antiguos: Aristóteles escribe sobre ellos, y es posible que los fósiles de un dinosaurio con pico, Protoceratops, estén detrás del mitológico grifo de los antiguos griegos. Pero el verdadero significado de los fósiles no se comenzó a valorar hasta mucho más tarde. Aun en pleno siglo XIX se explicaban como productos de fuerzas sobrenaturales, como organismos enterrados por el gran diluvio universal o como restos de especies vivas que habitaban en lugares remotos e inexplorados.
En estos restos petrificados, sin embargo, se esconde la historia de la vida. Pero ¿cómo podemos descifrarla? Para empezar, como es lógico, hay que tener los fósiles, y muchos. Luego hay que ponerlos en el orden correcto, de más antiguo a más reciente. Y luego hay que averiguar exactamente cuándo se formaron. Cada uno de estos requisitos conlleva sus propios desafíos.
Una vez bien enterradas en el sedimento, las partes duras de los fósiles van siendo infiltradas o reemplazadas por minerales disueltos. El resultado es un molde de un organismo vivo que queda comprimido en una roca por la presión de unos sedimentos que no dejan de acumularse encima. El hecho de que las partes blandas de plantas y animales no se fosilicen fácilmente crea de inmediato un sesgo importante en nuestro conocimiento de las especies antiguas. Los huesos y los dientes son abundantes, igual que las conchas y los esqueletos externos de insectos y crustáceos. Pero los gusanos, las medusas, las bacterias y los organismos frágiles como las aves son mucho más escasos, lo mismo que las especies terrestres por comparación con las acuáticas. Durante el primer 80 por 100 de la historia de la vida, todas las especies eran de cuerpo blando, así que para observar las fases más primitivas e interesantes de la evolución disponemos sólo de una ventana empañada, y para ver el origen de la vida, de ninguna.
Una vez formado, un fósil tiene que sobrevivir a los continuos procesos de desplazamiento, plegamiento, calentamiento y aplastamiento de las corteza de la Tierra, que acaban por destruir la mayoría de los fósiles. Luego hay que descubrirlo. La mayoría, enterrados como están a gran profundidad bajo la superficie de la Tierra, nos resultan inaccesibles. Sólo cuando los sedimentos son levantados y quedan expuestos gracias a la erosión por el viento o la lluvia, pueden ser atacados por el martillo del paleontólogo. Además, sólo hay una pequeña ventana de tiempo antes de que estos fósiles parcialmente expuestos acaben también desgastados y borrados por la acción del viento, el agua y otros agentes de la meteorología.
Si se tienen en cuenta todos estos requisitos, está claro que el registro fósil tiene que ser incompleto. Pero ¿en qué medida? El número total de especies que en algún momento han vivido en la Tierra se estima entre 17 millones (lo que probablemente sea una drástica subestima puesto que en la actualidad viven al menos 10 millones de especies) y 4.000 millones. Dado que hemos descubierto alrededor de 250.000 especies distintas de fósiles, podemos estimar que nuestro registro fósil apenas corresponde a entre 0,1 y 1 por 100 de todas las especies, ¡una muestra muy deficiente de la historia de la vida! Deben de haber existido muchas especies sorprendentes que hemos perdido para siempre. Con todo, disponemos de fósiles suficientes para hacernos una buena idea de cómo se produjo la evolución y para discernir cómo los grandes grupos se fueron escindiendo unos de otros.
Irónicamente, quienes primero pusieron orden en el registro fósil no fueron evolucionistas sino geólogos que además eran creacionistas y aceptaban la explicación de la vida que ofrece el libro del Génesis. Estos primeros geólogos se limitaron a ordenar las distintas capas de rocas que iban descubriendo (a menudo durante las excavaciones de canales que acompañaron a la revolución industrial inglesa) con arreglo a unos principios fundamentados de sentido común. Como los fósiles aparecen en rocas sedimentarias que comenzaron como limos en océanos, ríos o lagos (o, más raramente, como dunas de arena o depósitos glaciales), las capas o «estratos» más profundos tenían que haberse depositado antes que los más cercanos a la superficie. Las rocas más jóvenes descansan sobre rocas más antiguas. Pero no en todos los lugares se depositaron todas las capas: no siempre en un mismo lugar había agua para formar sedimentos.
Por consiguiente, para establecer el orden completo de los estratos era necesario correlacionar los hallados en distintas localidades de todo el mundo. Si una capa del mismo tipo de roca y con el mismo tipo de fósiles aparece en dos lugares distintos, es razonable suponer que la capa es de la misma edad en ambas localidades. Por ejemplo, si encontramos cuatro capas de roca en un lugar (que podemos llamar, de la más superficial a la más profunda, ABDE), y luego encontramos tan sólo dos de esas capas en otro lugar, pero con una nueva capa entre las dos, por ejemplo BCD, podemos inferir que este registro incluye al menos cinco capas de roca que, de más recientes a más antiguas, serían ABCDE. Este principio de superposición lo ideó inicialmente en el siglo XVII el polímata danés Nicolaus Steno, quien más adelante se convertiría en arzobispo y fue beatificado por el papa Juan Pablo II en 1988 en lo que seguramente constituye el único caso de un beato que haya realizado una contribución científica importante. Con la ayuda del principio de Steno, el registro geológico se fue ordenando mediante un trabajo minucioso a lo largo de los siglos XVII y XVIII, desde las antiquísimas rocas del Cámbrico hasta las más recientes. Hasta aquí, bien. Pero esto sólo nos dice las edades relativas de las rocas, no sus edades reales o absolutas.
Desde aproximadamente 1945 podemos medir las edades reales de algunas rocas usando la radiactividad. Ciertos elementos radiactivos («radioisótopos») quedan incorporados en las rocas ígneas cuando se cristalizan a partir de la roca fundida bajo la superficie de la Tierra. Los radioisótopos se desintegran de manera gradual en otros elementos de acuerdo con una tasa constante, que suele expresarse en forma de una «vida media», es decir, el tiempo necesario para que desaparezca la mitad de un isótopo. Una vez conocida la vida media de un isótopo, en qué cantidad se encontraba en el momento de la formación de la roca (algo que los geólogos pueden determinar con precisión) y cuánto queda en la actualidad, es relativamente sencillo estimar la edad de la roca. Cada isótopo se desintegra con una tasa característica. Las rocas antiguas suelen datarse con uranio-238 (U-238), que se encuentra en un mineral común, el circón. El U-238 tiene una vida media de unos 700 millones de años. El carbono-14, con una vida media de 5.730 años, se utiliza en rocas mucho más recientes, o incluso en artefactos humanos como los manuscritos del mar Muerto. Es frecuente que en un mismo material se encuentren varios radioisótopos, de modo que podemos cotejar las dataciones, y la edades invariablemente concuerdan. Las rocas que contienen fósiles, sin embargo, no son ígneas, sino sedimentarias, y no es posible datarlas de manera directa. Pero podemos obtener las edades de los lechos fosilíferos como el intervalo de edades de las capas de rocas ígneas adyacentes que contengan radioisótopos.
Los oponentes de la evolución a menudo atacan la fiabilidad de estas dataciones aduciendo que las tasas de desintegración radiactiva podrían haber cambiado con el tiempo o con las presiones físicas que hayan experimentado las rocas. Esta objeción suelen plantearla los creacionistas de la «tierra joven», quienes sostienen que nuestro planeta no tiene más de seis a diez mil años de edad. Pero su objeción es engañosa. Como los distintos radioisótopos de una roca se desintegran de forma distinta, si sus tasas de desintegración cambiaran no nos darían dataciones concordantes. Además, las vidas medias de los isótopos no cambian cuando los científicos los someten a temperaturas y presiones extremas en el laboratorio. Y en aquellos casos en que las fechas radiométricas han podido compararse con fechas del registro histórico, como ocurre con el método del carbono-14, concuerdan de manera invariable. Es la datación radiométrica de los meteoritos lo que nos dice que la Tierra y el sistema solar tienen 4.600 millones de años. (Las rocas más antiguas de la Tierra son algo más jóvenes, de unos 4.300 millones de años en muestras del norte de Canadá, porque las más antiguas han quedado destruidas por movimientos de la corteza terrestre.)
Hay aún otras maneras de contrastar la exactitud de la datación radiométrica. Una de ellas consiste en utilizar la biología, como hizo John Well, de la Universidad de Cornell, en un ingenioso estudio de unos corales fósiles. La datación con radioisótopos había mostrado que estos corales habían vivido durante el período Devónico, hace unos 380 millones de años. Pero Well logró también hallar la edad de estos corales con sólo mirarlos detenidamente. Lo que hizo fue aprovechar el hecho de que, con el tiempo, la fricción producida por las mareas va frenando gradualmente la rotación de la Tierra. Cada día, una revolución de la Tierra es un poquito más larga que el día anterior. Nada que podamos notar; para ser precisos, la longitud de un día se incrementa en aproximadamente dos segundos cada 100.000 años. Como la duración de un año (el tiempo que tarda la Tierra en dar la vuelta al Sol) no cambia con el tiempo, esto significa que el número de días por año debe disminuir con el tiempo. A partir de la tasa de frenado, Wells calculó que cuando sus corales estaban vivos (hace 380 millones de años, si la datación radiométrica era correcta) cada año debía de contener unos 396 días, cada uno de ellos de 22 horas de duración. Si hubiera algún modo de que los propios fósiles nos dijeran cuál era la longitud del día cuando vivieron, podríamos comprobar si esa longitud concuerda con las 22 horas que predice la datación radiométrica.
Lo bueno del caso es que los corales pueden decirnos eso, pues a medida que crecen dejan registrado en su cuerpo el número de días del año. Los corales vivos producen anillos de crecimiento diarios y anuales. En los especímenes fósiles, podemos ver cuántos anillos diarios separan cada anillo anual; dicho de otro modo, cuántos días había en un año cuando vivían aquellos corales. Si conocemos la tasa de frenado producida por las mareas, podemos comparar la edad «mareal» con la edad «radiométrica». Después de contar los anillos en sus corales devónicos, Wells encontró que habían experimentado unos 400 días por año, lo que corresponde a un día de 21,9 horas, una desviación minúscula de la predicción de 22 horas. Esta ingeniosa calibración biológica refuerza nuestra confianza en la exactitud de la datación radiométrica.
Los hechos
¿Qué podemos considerar pruebas de la evolución en el registro fósil? Hay varios tipos. En primer lugar, la visión global de la evolución: un examen de la secuencia entera de estratos debería mostramos que las primeras formas de vida eran bastante sencillas, y que las formas más complejas fueron apareciendo con el tiempo. Además, los fósiles más recientes deberían ser los más parecidos a las especies vivas en la actualidad.
También deberíamos hallar ejemplos del cambio evolutivo dentro de un mismo linaje, es decir, cambios a lo largo del tiempo en una especie de animal o de planta. Las especies más tardías deberían poseer caracteres que hagan que se parezcan a descendientes de las especies anteriores. Y como la historia de la vida implica la escisión de especies a partir de antepasados comunes, también deberíamos ser capaces de ver esta escisión en el registro fósil, y de encontrar indicios de los antecesores comunes. Por ejemplo, los anatomistas del siglo XIX predijeron, a partir de sus semejanzas corporales, que los mamíferos habían evolucionado de antiguos reptiles. Por lo tanto, deberíamos poder encontrar fósiles de reptiles en una secuencia temporal que los acerque cada vez más a los mamíferos. Naturalmente, como el registro fósil es incompleto, no cabe esperar que podamos documentar todas y cada una de las transiciones entre las principales formas de vida. Pero al menos deberíamos encontrar algunas.
En El origen, Darwin se lamenta de lo fragmentario del registro fósil. Por aquel entonces carecíamos de series transicionales de fósiles o «eslabones perdidos» entre formas principales que sirvieran para documentar el cambio evolutivo. Algunos grupos, como las ballenas, aparecían de repente en el registro, sin antepasados conocidos. Pero aun así Darwin disponía de algunos indicios fósiles a favor de la evolución, entre ellos la observación de que las plantas y animales antiguos eran muy distintos de las especies actuales, y que se parecían más a éstas cuanto más recientes fueran las rocas donde se habían descubierto. Observó también que los fósiles de las capas adyacentes eran más parecidos entre sí que los hallados en capas más separadas, lo que implicaba un proceso continuo y gradual de divergencia. Más aún, en un lugar dado, los fósiles de los lechos de roca depositados más recientemente tendían a parecerse a las especies modernas que vivían en la zona, y no a las especies de otras partes del mundo. Por ejemplo, los marsupiales se encontraban en profusión solo en Australia, que es donde viven la mayoría de los marsupiales actuales. Esto sugería que las especies modernas descendían de las fósiles. (Entre los marsupiales fósiles se encuentran algunos de los mamíferos más extraños que hayan vivido nunca, entre ellos un canguro gigante de unos tres metros con el rostro plano, enormes garras y un solo dedo en cada pie.)
Lo que no tenía Darwin eran fósiles suficientes para aportar evidencias incontestables de cambios graduales en especies, o de antepasados comunes. Pero desde entonces los paleontólogos han descubiertos muchísimos fósiles que satisfacen las predicciones mencionadas anteriormente. Hoy podemos demostrar cambios continuos dentro de linajes de animales; disponemos de abundantes indicios de antepasados comunes y formas transicionales (han aparecido, por ejemplo, los antepasados perdidos de las ballenas); y hemos excavado lo bastante hondo como para ver los inicios de la vida compleja.
Grandes patrones
Ahora que tenemos todos los estratos en orden y hemos estimado sus edades, podemos leer el registro fósil de abajo arriba. La Figura 3 presenta una línea de tiempo simplificada de la historia de la vida en la que se muestran los principales eventos biológicos y geológicos que se han producido desde la aparición de los primeros organismos, hace unos 3.500 millones de años.[6] Este registro ofrece una imagen inequívoca del cambio desde lo más simple a lo más complejo. Aunque la figura muestra las «primeras apariciones» de grupos como los reptiles y los mamíferos, no debe interpretarse que las formas modernas aparezcan en el registro fósil de repente, como si salieran de la nada. Bien al contrario, en la mayoría de los grupos vemos una evolución gradual a partir de formas más primitivas (las aves y los mamíferos, por ejemplo, evolucionaron a lo largo de millones de años a partir de antepasados reptilianos). La existencia de transiciones graduales entre grandes grupos, tal como se comenta más adelante, significa que la asignación de una fecha de «primera aparición» es un tanto arbitraria.
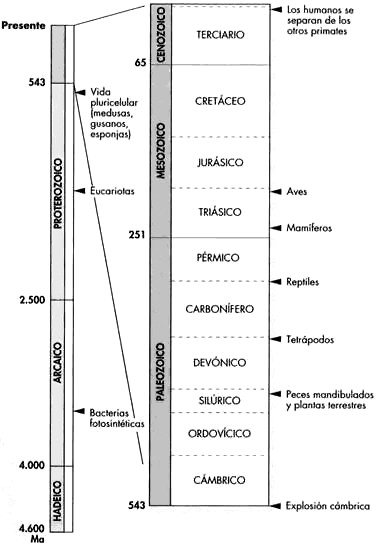
FIGURA 3. Registro fósil con indicación de la primera aparición de las diversas formas de vida que surgieron desde la formación de la Tierra hace unos 4.600 millones de años (Ma). Nótese que la vida pluricelular se originó y diversificó sólo en el último 15 por 100 de la historia de la vida. Los grupos aparecen en escena de un modo ordenado de acuerdo con su evolución, y muchos aparecen después de transiciones fósiles conocidas desde sus antepasados.
Los primeros organismos, unas simples bacterias fotosintéticas, aparecen en los sedimentos de hace unos 3.500 millones de años, tan sólo unos 1.000 millones de años después de la formación del planeta. Estos organismos unicelulares fueron lo único que ocupó la Tierra durante los 2.000 millones de años siguientes, tras los cuales vemos la aparición de los primeros y sencillos «eucariotas», los organismos con células verdaderas dotadas de núcleo y cromosomas. Más tarde, hace unos 600 millones de años, surge el abanico completo de organismos relativamente simples pero pluricelulares, como los gusanos, las medusas y las esponjas. Estos grupos se diversificaron durante varios millones de años; las plantas y los tetrápodos terrestres (animales de cuatro patas, los primeros de los cuales fueron peces de aletas lobuladas) no aparecieron hasta hace unos 400 millones de años. Naturalmente, muchos de los grupos que aparecieron más tempranamente persistieron: las bacterias fotosintéticas, las esponjas y los gusanos aparecen en el registro fósil más primitivo pero todavía siguen con nosotros.
Unos 50 millones de años más tarde encontramos los primeros anfibios verdaderos, y tras otros 50 millones de años, los reptiles. Los primeros mamíferos no hacen acto de presencia hasta hace unos 250 millones de años (a partir de antepasados reptiles, como se había predicho), y las primeras aves, descendientes también de reptiles, aparecen unos 50 millones de años más tarde. Tras la aparición de los primeros mamíferos, éstos, junto con los insectos y las plantas terrestres, se van diversificando cada vez más, y a medida que nos acercamos a las rocas más superficiales, los fósiles se parecen cada vez más a las especies vivas. Los humanos son unos recién llegados a este escenario: nuestro linaje se separa del de otros primates hace tan sólo unos 7 millones de años, apenas una finísima rebanada del tiempo entero de la evolución. Se ha propuesto toda una serie de imaginativas analogías para resaltar lo ultimísimo de nuestra llegada, y merece la pena hacerlo una vez más. Si el recorrido entero de la evolución se comprimiera en un solo año, las primeras bacterias habrían aparecido a finales de marzo, pero no veríamos los primeros antepasados de los humanos hasta las seis de la mañana del 31 de diciembre. La edad dorada de Grecia, alrededor del 500 a. C., se produciría a tan sólo treinta segundos de la medianoche.
Aunque el registro fósil de las plantas es más escaso, pues carecen de partes duras que se fosilicen fácilmente, su patrón evolutivo es muy parecido. Las más antiguas son musgos y algas, seguidas de la aparición de los helechos, y éstos de las coníferas, los árboles de hoja caduca y, por último, las plantas con flor.
Así que la aparición de las especies a lo largo del tiempo, tal como se puede ver en los fósiles, está muy lejos de ser aleatoria. Los organismos simples evolucionaron antes que los complejos, los antepasados que se habían predicho, antes que sus descendientes. Los fósiles más recientes son los más parecidos a las especies vivas. Y disponemos de fósiles transicionales que conectan los principales grupos. Ninguna teoría de la creación especial, ninguna teoría que no sea la evolución, puede explicar estos patrones.
La evolución y la especiación en los fósiles
Para poner de manifiesto el cambio evolutivo gradual dentro de un mismo linaje se necesita una buena secuencia de sedimentos, con preferencia que se hayan depositado con rapidez (de manera que cada período de tiempo venga representado por un lecho grueso de sedimentos y sea más fácil observar los cambios), y en la que no falten capas (la falta de una capa en medio de una secuencia hace que una transición evolutiva suave parezca un «salto» brusco).
Los animales marinos de pequeño tamaño, como los del plancton, son ideales para este propósito. Son extraordinariamente abundantes, muchos tienen partes duras y, lo mejor de todo, caen directamente al fondo del océano al morir, donde se acumulan en capas que forman secuencias continuas. Obtener muestras ordenadas de estas capas es fácil: basta con clavar un largo tubo en el fondo del mar, extraer con él una columna de sedimento (un testigo), y leerlo (datarlo) desde el fondo hasta la superficie.
Cuando se sigue una misma especie de fósil a lo largo del testigo, a menudo puede verse cómo evoluciona. La Figura 4 muestra un ejemplo de evolución en un diminuto protozoo unicelular marino que construye una concha en espiral y crea nuevas cámaras a medida que crece. Estas muestras provienen de secciones de un testigo de doscientos metros de longitud obtenido en los sedimentos del fondo marino cerca de la costa de Nueva Zelanda. La figura muestra cómo cambia con el tiempo un solo carácter: el número de cámaras de la última espiral de la concha. Lo que vemos es un cambio bastante suave y gradual en el tiempo desde una media de 4,8 cámaras por espiral al principio de la secuencia hasta 3,3 cámaras al final, una reducción de alrededor del 30 por 100.
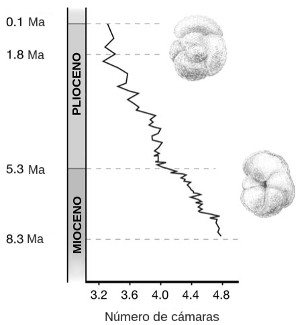
FIGURA 4. Registro de fósiles preservados en un testigo de sedimento marino que muestra los cambios evolutivos en un foraminífero marino, Globorotalia conoidea, a lo largo de 8 millones de años. La escala indica el número de cámaras de la última espiral de la concha, promediado entre todos los especímenes contados en cada sección del testigo.
La evolución, aunque gradual, no tiene por qué proceder siempre de forma suave o con un ritmo constante. La Figura 5 muestra un cambio más irregular en otro microorganismo marino, el radiolario Pseudocubus vema. En este caso los geólogos tomaron muestras regularmente espaciadas de un testigo de dieciocho metros extraído cerca de la Antártida, correspondiente a unos 2 millones de años de sedimentos. El carácter que midieron fue la anchura de la base cilíndrica del animal (su «tórax»). Aunque el tamaño aumenta en un 50 por 100 hacia el final de la serie, no sigue una tendencia suave. Hay períodos en los que el tamaño apenas cambia, intercalados por períodos de cambio más rápido. Esta pauta es bastante común en los fósiles, y puede entenderse completamente si los cambios que observamos fueron impulsados por factores ambientales, por ejemplo fluctuaciones en el clima o la salinidad. El entorno cambia de forma esporádica e irregular, y con él aumenta y disminuye la presión de la selección natural.
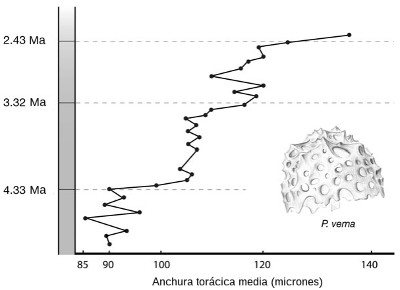
FIGURA 5. Cambios evolutivos en la anchura torácica del radiolario Pseudocubus vema durante un período de 2 millones de años. Los valores son promedios poblacionales para cada sección del testigo.
Examinemos ahora la evolución de unos organismos más complejos: los trilobites. Los trilobites eran artrópodos, el mismo grupo al que pertenecen los insectos y las arañas. Como estaban protegidos por una cubierta dura, son muy comunes en las rocas antiguas (seguro que los tienen a la venta en algún museo cercano). Peter Sheldon, entonces en el Trinity College de Dublín, recogió fósiles de trilobites de un lecho de esquisto de Gales que comprendía un período de unos 3 millones de años. Encontró en esta roca ocho linajes distintos de trilobites, y todos mostraron con el tiempo cambios evolutivos en el número de «costillas pigidiales», los segmentos de la última sección del cuerpo. La Figura 6 muestra los cambios en algunos de estos linajes. Aunque a lo largo de todo el período muestreado todas las especies presentaron un aumento neto del número de segmentos, los cambios entre las distintas especies no sólo no corren parejos, sino que a veces van en direcciones distintas durante el mismo período.
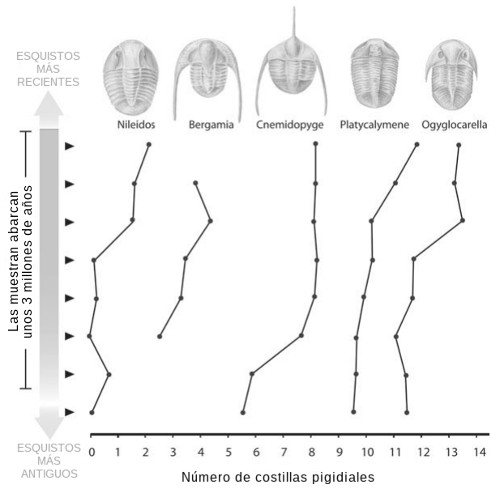
FIGURA 6. Cambios evolutivos en el número de «costillas pigidiales» (segmentos de la sección posterior) de cinco grupos de trilobites del Ordovícico. Los valores corresponden a la media de la población para cada sección de una muestra de esquisto de hace 3 millones de años. Las cinco especies (y otras tres que no se muestran aquí) presentan un cambio neto en el número de costillas a lo largo del período, y sugieren que la selección natural actuó a largo plazo, pero que las especies no cambiaron perfectamente en paralelo.
Lamentablemente, no tenemos la menor idea de cuáles fueron las presiones selectivas que impulsaron los cambios evolutivos en estas especies del plancton y de trilobites. Siempre es más fácil documentar la evolución en el registro fósil que entender su causa, pues se preservan los fósiles, pero no el medio donde habitaban. Lo que podemos decir es que hubo evolución, que fue gradual y que varió tanto en el ritmo como en la dirección.
Además de ilustrar la evolución dentro de un linaje, el plancton marino también nos ofrece ejemplos de la escisión de un linaje. La Figura 7 muestra una especie ancestral de plancton que se divide en dos descendientes que se distinguen por el tamaño y la forma del cuerpo. Lo interesante es que la nueva especie, Eucyrtidium matuyamai, evolucionó al principio en un área al norte de la zona donde se tomaron estos testigos de sedimento, y más tarde invadió el área donde habitaba su antepasado. Como veremos en el capítulo 7, la formación de una especie nueva a menudo comienza con el aislamiento geográfico de dos poblaciones de una especie.
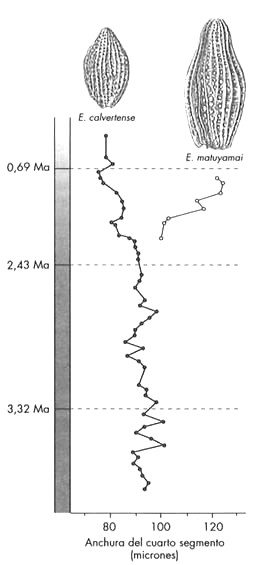
FIGURA 7. Evolución y especiación en dos especies del radiolario planctónico Eucyrtidium, a partir de un testigo de sedimento que comprende más de 3,5 millones de años. Los puntos, que representan la anchura del cuarto segmento, corresponden al promedio para cada especie y sección del testigo. En las áreas al norte de donde se extrajo este testigo, una población ancestral de E. calvertense se fue haciendo mayor, adquiriendo gradualmente el nombre de E. matuyamai al alcanzar un mayor tamaño. E. matuyamai invadió después el área de distribución de su pariente, tal como muestra el gráfico, y ambas especies, al habitar ahora en el mismo lugar, comenzaron a divergir en el tamaño del cuerpo. Esta divergencia podría explicarse por la actuación de la selección natural para reducir la competencia por el alimento entre las dos especies.
Hay centenares de ejemplos más de cambios evolutivos en fósiles, tanto graduales como puntuados, en especies de grupos tan distintos como los moluscos, los roedores y los primates. Y hay ejemplos también de especies que apenas han cambiado. (¡La teoría de la evolución no dice que todas las especies tengan que cambiar!) Pero una lista de estos casos no modificaría el mensaje: el registro fósil no ofrece indicio alguno a favor de la predicción creacionista de que todas las especies aparecieron de un solo golpe y permanecieron inmutables desde entonces. Por el contrario, las formas de vida aparecen en el registro de acuerdo con una secuencia evolutiva, evolucionan y divergen.
«Eslabones perdidos»
Los cambios en las especies marinas nos ofrecen indicios de la evolución, pero ésta no es la única lección que nos enseña el registro fósil. Lo que de verdad apasiona a la gente, y sobre todo a los biólogos y paleontólogos, son las formas transicionales, los fósiles que cierran la brecha entre dos tipos muy distintos de organismos vivos. ¿Es realmente cierto que las aves vienen de unos reptiles, los animales terrestres de unos peces y las ballenas de unos animales terrestres? Si es así, ¿dónde están los indicios fósiles que lo avalan? Algunos creacionistas admiten la posibilidad de que con el tiempo se produzcan pequeños cambios en tamaño y forma —un proceso denominado microevolución—, pero rechazan la idea de que a partir de un tipo de animal o planta pueda aparecer otro tipo muy distinto (macroevolución). Los defensores del diseño inteligente sostienen que este tipo de diferencia requiere la intervención directa de un creador.[7] Aunque en El origen Darwin no pudo aportar como pruebas formas de transición, hoy estaría muy satisfecho de ver cómo su teoría ha sido confirmada por los frutos de la moderna paleontología. Entre éstos se incluyen numerosas especies cuya existencia se había predicho hace muchos años, pero que sólo se han desenterrado durante las últimas décadas.
Pero ¿qué puede considerarse indicio fósil de una transición macroevolutiva? De acuerdo con la teoría de la evolución, para cada par de especies, por distintas que sean, hubo en algún momento una única especie que fue antepasado de las dos. A esta especie podemos denominarla «eslabón perdido». Como hemos visto, la probabilidad de descubrir esta especie ancestral en el registro fósil es casi nula. El registro fósil es, sencillamente, demasiado fragmentario.
Pero no tenemos que rendirnos, porque podemos encontrar alguna otra especie en el registro fósil que sea un pariente próximo del verdadero «eslabón perdido» y sirva a la perfección para documentar un antepasado común. Veamos un ejemplo. Ya en tiempos de Darwin los biólogos conjeturaron, a partir de observaciones anatómicas como las similitudes en la estructura del corazón y del cráneo, que las aves eran parientes cercanos de los reptiles. Especularon entonces que debía existir un antepasado común que, por medio de un proceso de especiación, habría producido dos linajes, uno de los cuales acabaría dando todas las aves modernas y el otro todos los reptiles actuales.
¿Qué aspecto debía de tener este antepasado común? Nuestra intuición nos dice que debía de estar a medio camino entre los reptiles modernos y las aves modernas, con una mezcla de caracteres de ambos tipos de animal. Pero no tenía por qué ser así, como Darwin decía con claridad en El origen:
Considerando dos especies cualesquiera, he encontrado difícil evitar el imaginarse formas directamente intermedias entre ellas; pero ésta es una opinión errónea; hemos de buscar siempre formas intermedias entre cada una de las especies y un antepasado común y desconocido, y este antepasado, por lo general, habrá diferido en algunos conceptos de todos sus descendientes modificados.
Como los reptiles aparecen en el registro fósil antes que las aves, podemos aventurar que el antepasado común de aves y reptiles fue un antiguo reptil y tendría aspecto de tal. Hoy sabemos que ese antepasado común era un dinosaurio. Su aspecto general daría pocas pistas de que era en realidad un «eslabón perdido», de que uno de los linajes de su descendencia acabaría dando origen con el tiempo a todas las aves modernas, y el otro a más dinosaurios. Los caracteres genuinos de las aves, como las alas y la gran quilla para el anclaje de los músculos del vuelo, sólo habrían evolucionado más tarde en la rama que condujo a las aves. Y a medida que este linaje progresaba de los reptiles a las aves, habría dado origen a muchas especies con mezclas de caracteres de reptiles y de aves. Algunas de esas especies se extinguieron; otras siguieron evolucionando hasta llegar a lo que hoy conocemos como las aves modernas. Es en estos grupos de antiguas especies, en los parientes de especies cerca de los puntos de ramificación, donde debemos buscar los indicios de los ancestros comunes.
Demostrar la ascendencia común de dos grupos no exige, por consiguiente, que hallemos fósiles de la especie exacta que fue su antepasado común, ni siquiera de especies situadas en la línea directa de descendencia desde el antepasado al descendiente. Basta con que descubramos fósiles que posean los tipos de caracteres que vinculan a los dos grupos, y, lo que es igualmente importante, que la datación de los fósiles corresponda al momento correcto del registro geológico. Una «especie transicional» no es lo mismo que una «especie ancestral»; es simplemente una especie que presenta una mezcla de los caracteres de organismos que vivieron antes y después que ella. Habida cuenta de la naturaleza fragmentaria del registro fósil, hallar estas formas en los momentos adecuados del registro es un objetivo razonable y realista. En la transición de reptil a ave, por ejemplo, las formas transicionales deberían tener el aspecto de reptiles primitivos, pero con algunos caracteres propios de las aves. Y deberíamos encontrar estas formas de transición después de que hayan evolucionado los reptiles, pero antes de que aparezcan las aves modernas. Además, las formas transicionales no tienen por qué estar en la línea directa de descendencia desde un antepasado a un descendiente vivo en la actualidad, sino que pueden ser primos evolutivos que hayan acabado extinguiéndose. Como veremos, los dinosaurios que dieron origen a las aves tenían plumas, pero persistieron algunos dinosaurios con plumas cuando ya habían evolucionado organismos con un aspecto más parecido al de las aves actuales. Estos dinosaurios con plumas de épocas posteriores todavía constituyen indicios de la evolución, porque nos dicen algo acerca de dónde vienen las aves.
La datación y, hasta cierto punto, la apariencia física de las formas de transición pueden predecirse a partir de la teoría de la evolución. Algunas de las predicciones más recientes y vistosas que se han podido satisfacer tienen que ver con nuestro propio grupo: los vertebrados.
A la tierra firme: de los peces a los anfibios
Una de las predicciones mejor satisfechas de la biología evolutiva es el descubrimiento, en 2004, de una forma de transición entre los peces y los anfibios. Se trata del fósil de la especie Tiktaalik roseae, que nos dice mucho sobre el paso de los vertebrados a la tierra firme. Su descubrimiento es una vindicación rotunda de la teoría de la evolución.
Hasta hace unos 390 millones de años, los peces eran los únicos vertebrados. Pero, 30 millones de años más tarde, encontramos animales que son claramente tetrápodos, vertebrados de cuatro patas que caminaban por la tierra. Estos primitivos tetrápodos eran en muchos sentidos como los modernos anfibios: tenían la cabeza y el cuerpo planos, un cuello bien formado y extremidades bien desarrolladas, incluidas las cinturas escapular y pélvica. Pero presentan asimismo caracteres que los vinculan claramente con los peces primitivos, en particular con el grupo conocido como «peces de aletas lobuladas», así llamados a causa de sus grandes aletas óseas que les permitían apoyarse en el fondo de las lagunas y ríos someros. Entre las estructuras propias de los peces presentes en los primeros tetrápodos encontramos las escamas, los huesos de las extremidades y los huesos del cráneo (Figura 8).
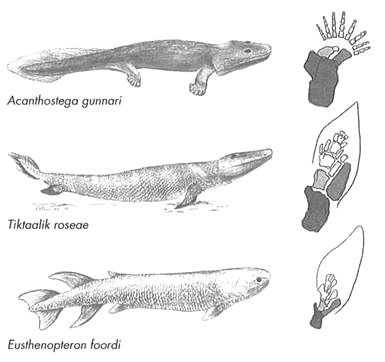
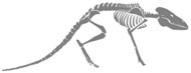
FIGURA 8. Invasión de la tierra firme. Un primitivo pez lobulado (Eusthenopteron foordi) de hace unos 385 millones de años; un tetrápodo terrestre (Acanthostega gunnari) de Groenlandia, de hace unos 365 millones de años; y la forma de transición, Tiktaalik roseae, de la isla Ellesmere, de hace unos 375 millones de años. El carácter intermedio de la forma corporal de Tiktaalik es soslayado por el carácter intermedio de sus extremidades, que tienen una estructura ósea a medio camino entre la de las robustas aletas del pez de aletas lobuladas y las extremidades ambulatorias, todavía más robustas, del tetrápodo. Los huesos que evolucionaron hacia los huesos de los brazos y las piernas de los mamíferos modernos aparecen en color gris; el hueso con el tono más oscuro se convirtió en nuestro húmero, y los de tono gris medio y claro se convirtieron en el radio y el cubito, respectivamente.
¿Cómo evolucionaron los peces primitivos para poder sobrevivir en la tierra? Ésta es la pregunta que interesaba, o más bien obsesionaba, a mi colega de la Universidad de Chicago, Neil Shubin. Neil había dedicado años al estudio de la evolución de las extremidades a partir de las aletas, y estaba decidido a entender los primeros estadios de esa evolución.
Aquí es donde entra la predicción. Si hace 390 millones de años había peces de aletas lobuladas pero no vertebrados terrestres, y hace 360 millones de años ya había vertebrados claramente terrestres, ¿dónde podían encontrarse las formas transicionales? En algún lugar a medio camino. Siguiendo esta lógica, Shubin predijo que si habían existido formas transicionales, sus fósiles deberían encontrarse en estratos de hace unos 375 millones de años. Además, las rocas deberían tener su origen en aguas dulces, no en sedimentos marinos, porque tanto los peces de aletas lobuladas posteriores como los anfibios habitaban las aguas dulces.
Shubin y sus colegas buscaron entonces en sus libros de geología un mapa de los afloramientos de rocas sedimentarias de agua dulce de la edad apropiada, y decidieron centrar sus esfuerzos en una región paleontológicamente inexplorada del Ártico canadiense: la isla Ellesmere, que se encuentra en el océano Ártico, al norte de Canadá. Tras cinco años de costosa e infructuosa búsqueda, por fin dieron con un filón: un grupo de esqueletos fósiles apilados en una roca sedimentaria formada en un antiguo río. La primera vez que Shubin vio el rostro del fósil asomando por la roca, supo que por fin había encontrado su forma transicional. En honor al pueblo inuit y al patrocinador que ayudó a costear las expediciones, el fósil recibió el nombre de Tiktaalik roseae (Tiktaalik significa «pez grande de agua dulce» en inuit, y roseae es una referencia críptica al patrocinador anónimo).
Tiktaalik posee caracteres que lo convierten en un vínculo directo entre los anteriores peces de aletas lobuladas y los anfibios posteriores (Figura 8). Con agallas, escamas y aletas, se trata claramente de un pez que vivía en el agua. Pero también posee características de anfibio. Para empezar, tiene la cabeza plana como la salamandra, con los ojos y las narinas en la parte superior y no a los lados del cráneo. Esto sugiere que vivía en aguas someras y podía ver, y probablemente respirar, por encima de la superficie. Las aletas son más robustas, lo que permitía al animal sostenerse sobre ellas y levantarse para explorar su entorno. Y, al igual que los primeros anfibios, Tiktaalik tiene cuello. Los peces carecen de cuello, pues tienen el cráneo unido directamente a las clavículas.
Lo más importante es, sin embargo, que Tiktaalik posee dos caracteres nuevos que habrían de ayudar a sus descendientes a invadir el medio terrestre. El primero es un conjunto de costillas robustas que ayudaban al animal a bombear el aire hasta sus pulmones y a mover el oxígeno desde las agallas (Tiktaalik podía respirar de las dos maneras). Y en lugar de los numerosos y diminutos huesos de las aletas de los peces de aletas lobuladas, Tiktaalik tenía en los miembros menos huesos, pero más robustos, huesos parecidos en número y posición a los de todos los animales terrestres que los siguieron, incluidos nosotros mismos. De hecho, sus miembros se pueden describir adecuadamente como mitad aleta, mitad pata.
No cabe duda de que Tiktaalik estaba bien adaptado a vivir y arrastrarse en aguas someras, a mirar por encima de la superficie del agua y a respirar aire. Conocida su estructura, podemos imaginar cuál debió de ser el siguiente paso evolutivo crucial, que con toda probabilidad comportó una conducta novedosa. Unos pocos descendientes de Tiktaalik tuvieron el arrojo de aventurarse fuera del agua aguantados por sus robustas aletas-patas, quizá para alcanzar otro río (como hacen en la actualidad los curiosos peces del fango tropicales), para evitar depredadores, o quizá para buscar comida entre los muchos insectos gigantes que ya había producido la evolución. Si aventurarse en tierra firme comportaba ventajas, la selección natural habría ido moldeando esos exploradores de peces a anfibios. Ese primer pasito en el fango resultó ser un gran salto para los vertebrados, pues con el tiempo habría de conducir a la evolución de todos los animales terrestres dotados de columna vertebral.
El propio Tiktaalik no estaba preparado para vivir fuera del agua. Para empezar, todavía no había evolucionado hasta el punto de disponer de miembros que le permitieran caminar. Y todavía tenía agallas internas para respirar bajo del agua. Así que podemos hacer otra predicción. En algún lugar, en sedimentos de agua dulce de hace unos 380 millones de años, encontraremos uno de los primeros animales terrestres con agallas reducidas y extremidades algo más robustas que las de Tiktaalik.
Tiktaalik pone de manifiesto que nuestros antepasados eran peces depredadores con la cabeza aplanada que vivían en las aguas someras de los ríos. Es un fósil que vincula a los peces y los anfibios de maravilla. E igualmente maravilloso es el hecho de que su descubrimiento no se hubiese ya anticipado, sino predicho que se produciría en rocas de cierta edad y de cierto lugar.
La mejor manera de vivir el drama de la evolución es ver los fósiles o, mejor aún, tenerlos en las manos. Mis estudiantes gozaron de esta oportunidad cuando Neil trajo un molde de Tiktaalik a la clase, lo hizo pasar entre los alumnos y les indicó por qué cumplía con los requisitos de una verdadera forma transicional. Ésta fue, para ellos, la más tangible evidencia de que la evolución era cierta. ¿Con qué frecuencia tenemos en las manos un trozo de historia evolutiva, y especialmente uno que podría haber sido de un antepasado distante?
Al aire: el origen de las aves
¿Para qué sirve media ala? Desde los tiempos de Darwin, esta pregunta no ha dejado de plantearse con la intención de proyectar dudas sobre la evolución y la selección natural. Los biólogos nos dicen que las aves evolucionaron a partir de reptiles primitivos, pero ¿cómo pudo un animal terrestre desarrollar por medio de la evolución la capacidad de volar? La selección natural, nos dicen los creacionistas, no puede explicar esta transición porque requiere estadios intermedios en los cuales los animales contarían sólo con unos rudimentos de alas que parecen más un engorro que una ventaja selectiva.
Pero si se piensa con un poco de detenimiento, no cuesta tanto imaginar estadios intermedios en la evolución del vuelo, estadios que podrían haber sido útiles para sus poseedores. El planeo es obviamente el primer paso. Un paso que, además, ha evolucionado de manera independiente muchas veces: en animales placentarios, en marsupiales e incluso en reptiles. Las ardillas voladoras se las arreglan muy bien planeando con unos repliegues de la piel que extienden a los lados del cuerpo, una buena manera de pasar de un árbol a otro para escapar a los depredadores o buscar nueces. Y luego está el aun más notable «lémur volador» o colugo del sureste asiático, que posee una impresionante membrana que se extiende desde la cabeza hasta la cola. Se ha visto a un colugo planear una distancia de unos 140 metros, la longitud de seis pistas de tenis, con una pérdida de altura de tan sólo ¡doce metros! No es difícil imaginar el siguiente paso de la evolución: el batido de unos miembros como los del colugo para producir el vuelo verdadero, como vemos en los murciélagos. Pero ya no tenemos que contentarnos con imaginar ese paso, pues tenemos fósiles que muestran con toda claridad cómo evolucionaron las aves voladoras.
Desde el siglo XIX, la similitud entre los esqueletos de las aves y de algunos dinosaurios llevó a los paleontólogos a teorizar que tenían un antepasado común; en concreto, los terópodos, unos ágiles dinosaurios carnívoros que caminaban sobre dos patas. El registro fósil de hace unos 200 millones de años contiene muchos terópodos, pero ninguno que parezca, ni siquiera vagamente, un ave. En estratos de hace unos 70 millones de años, encontramos fósiles con un aspecto bastante moderno. Si la evolución es cierta, entonces cabía esperar encontrar la transición de reptiles a aves en rocas de entre 70 y 200 millones de años.
Y allí estaban. El primer vínculo entre aves y reptiles llegó a conocerlo el propio Darwin, quien, curiosamente, apenas lo cita de pasada en las últimas ediciones de El origen, y sólo como una rareza. Se trata de la que quizá sea la más célebre de todas las formas de transición: Archaeopteryx lithographica, un animal del tamaño de un cuervo, que se descubrió en una cantera de caliza de Alemania en 1860. (El nombre Archaeopteryx significa «ala antigua» y lithographica hace referencia a la caliza de Solnhofen, de grano lo bastante fino como para usarse en la fabricación de placas litográficas, y también para preservar las impresiones de unas blandas plumas.) Archaeopteryx posee la combinación justa de caracteres que uno esperaría encontrar en una forma de transición. Y su edad, de unos 145 millones de años, la sitúa allí donde esperábamos encontrarla.
En realidad, Archaeopteryx es más un reptil que un ave. Su esqueleto es casi idéntico al de algunos dinosaurios terópodos. Así lo clasificaron algunos biólogos que, no habiendo examinado sus fósiles con el detenimiento suficiente, no vieron las plumas. (La Figura 9 muestra esta similitud entre los dos tipos.) Los caracteres reptilianos incluyen una mandíbula con dientes, una cola larga y ósea, garras, dedos separados en las alas (en las aves modernas estos huesos están fusionados, como puede verse al comer alitas de pollo) y un cuello unido al cráneo por detrás (como en los dinosaurios) en lugar de por la base (como en las aves modernas). Los caracteres de ave son sólo dos: largas plumas y un dedo oponible que probablemente servía para posarse en una percha. Todavía no está claro que este animal, aunque dotado de plumas, pudiera volar, aunque sus plumas asimétricas (con un lado más largo que el otro) sugieren que así es. Las plumas asimétricas, como las alas de los aviones, crean la forma de alerón necesaria para un vuelo aerodinámico. Pero aun en el caso de que pudiera volar, Archaeopteryx es sobre todo un dinosaurio. Es también lo que los evolucionistas llaman un «mosaico»: en lugar de tener todos sus caracteres a medio camino entre los reptiles y las aves, tiene algunos propios de las aves y la mayoría de los reptiles.
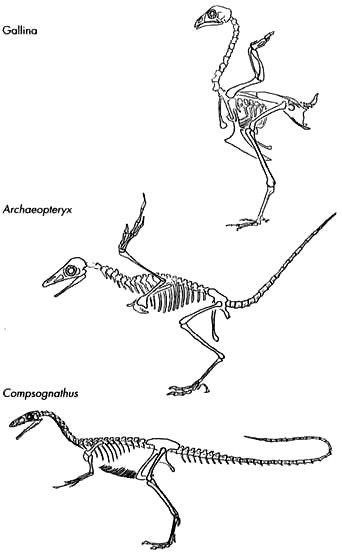
FIGURA 9. Esqueletos de un ave moderna (gallina), una forma de transición (Archaeopteryx) y un pequeño dinosaurio terópodo carnívoro y bípedo (Compsognathus), semejante a los antepasados de Archaeopteryx. Pueden apreciarse en Archaeopteryx algunos caracteres parecidos a los de las aves actuales (plumas y un dedo oponible), pero su esqueleto es muy similar al del dinosaurio, por ejemplo en los dientes, la pelvis reptiliana y la larga cola ósea. Archaeopteryx tenía más o menos el tamaño de un cuervo; Compsognathus era ligeramente mayor.
Tras el descubrimiento de Archaeopteryx, durante muchos años no se encontró ningún otro animal entre los reptiles y las aves, lo que dejaba abierta una amplia brecha entre las aves actuales y sus antepasados. Pero entonces, a mediados de la década de 1990, una avalancha de sorprendentes descubrimientos en China comenzó a cubrir la brecha. Estos fósiles, hallados en sedimentos lacustres que preservan las impresiones de las partes blandas, representan un verdadero desfile de dinosaurios terópodos con plumas.[8] Algunos presentan unas estructuras filamentosas muy pequeñas que les recubren todo el cuerpo y que probablemente sean unas primitivas plumas. Un fósil especialmente digno de mención es Sinomithosaurus millenii (Sinomithosaurus significa «ave-lagarto chino»), cuyo cuerpo aparece totalmente recubierto por unas plumas finas y alargadas, pero tan pequeñas que difícilmente podrían ayudarlo a volar (Figura 10A). Y sus garras, dientes y larga cola ósea muestran con toda claridad que este animal estaba muy lejos todavía de las aves modernas.[9] Otros dinosaurios presentan plumas de tamaño mediano en la cabeza y las extremidades anteriores. Incluso otros tienen grandes plumas en las extremidades anteriores y la cola, como las aves modernas. El más sorprendente de todos es Microraptor gui, el «dinosaurio de cuatro alas». A diferencia de las aves modernas, este extraño dinosaurio de unos 75 centímetros de longitud tenía cubiertas de plumas las extremidades anteriores y también las patas (Figura 10B), que una vez extendidas probablemente le sirvieran para planear.[10]
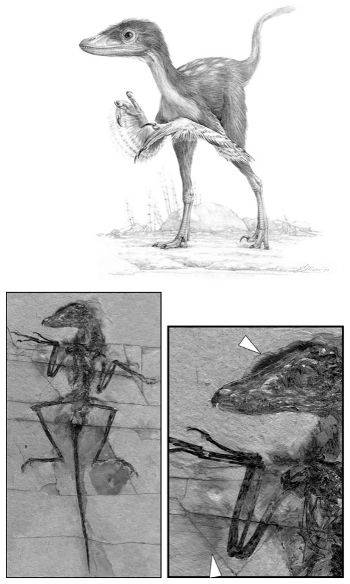
FIGURA 10A. El dinosaurio con plumas Sinomithosaurus millenii, fósil original de China (con una edad de unos 125 millones de años), y reconstrucción artística. El fósil muestra con claridad la impresión de plumas filamentosas, especialmente en la cabeza y las extremidades anteriores (flechas).

FIGURA 10B. El extraño dinosaurio «de cuatro alas» Microraptor gui, que tenía largas plumas en las cuatro extremidades. Estas plumas (flechas) se ven claramente en el fósil, de hace unos 120 millones de años. No está del todo claro si este animal podía volar o sólo planear, pero las «alas» posteriores casi con seguridad lo ayudaban a tomar tierra, tal como muestra el dibujo.
Por lo que podemos saber, los dinosaurios terópodos no sólo tenían caracteres que los acercaban a las aves: también se comportaban como aves. El paleontólogo norteamericano Mark Norell y su equipo han descrito dos fósiles que demuestran formas antiguas de comportamiento. Si de algún fósil puede decirse que es «conmovedor», es de éstos. Uno de ellos es un pequeño dinosaurio con plumas que duerme con la cabeza escondida bajo su antebrazo plegado como un ala, igual que hacen las aves modernas (Figura 11). El animal, que ha recibido el nombre científico de Mei long («dragón profundamente dormido», en chino), debió morir en medio del sopor. El otro fósil es una hembra de terópodo que encontró su fin mientras empollaba su puesta de huevos, lo que demuestra un comportamiento de cuidado de la nidada parecido al de las aves.

FIGURA 11. Comportamiento fósil: el dinosaurio terópodo con plumas Mei long (arriba) fosilizado en posición de percha, durmiendo con la cabeza escondida bajo la extremidad anterior. En medio: reconstrucción de Mei long a partir del fósil. Abajo: un ave actual (un gorrión joven) durmiendo en la misma posición.
Todos los fósiles de dinosaurios con plumas datan de hace entre 135 y 110 millones de años, más recientes que Archaeopteryx, que está datado en 145 millones de años. Esto significa que no podían ser antepasados directos de Archaeopteryx, pero podrían haber sido sus primos. Los dinosaurios con plumas probablemente continuaron existiendo después de que uno de los suyos diera origen a las aves. Por consiguiente, deberíamos poder descubrir dinosaurios con plumas más antiguos que pudieran ser antecesores de Archaeopteryx. El problema es que las plumas sólo se preservan en sedimentos especiales: los limos de grano fino de ambientes tranquilos como los fondos de lagos y lagunas. Y estas condiciones son muy poco frecuentes. Pero podemos hacer otra predicción evolutiva contrastable: algún día encontraremos fósiles de dinosaurio con plumas más antiguos que Archaeopteryx.[11]
No estamos seguros de si Archaeopteryx es la única especie que dio origen a todas las aves modernas. Parece improbable que sea el «eslabón perdido». Pero con independencia de ello, es un fósil de una larga serie (algunos descubiertos por el intrépido Paul Sereno) que documentan con toda claridad la aparición de las aves modernas. A medida que nos acercamos al presente vemos cómo la cola reptiliana se acorta, los dientes desaparecen, las garras se fusionan y aparece un gran hueso pectoral, la quilla, para el anclaje de los músculos del vuelo.
Cuando juntamos todas las piezas, vemos que los fósiles nos muestran que el plan esquelético básico de las aves, y las esenciales plumas, evolucionaron antes de que las aves pudieran volar. Hubo muchos dinosaurios con plumas, y éstas están claramente relacionadas con las de las aves modernas. Pero si las plumas no aparecieron como una adaptación para volar, ¿para qué demonios servían? No lo sabemos. Quizá como ornamentación o exhibición, para atraer a las parejas. Parece más probable, sin embargo, que sirvieran para aislar el cuerpo. A diferencia de los reptiles modernos, los terópodos podrían haber sido, aunque sólo parcialmente, animales de sangre caliente; y aunque no lo fueran, las plumas podrían haberlos ayudado a mantener la temperatura corporal. Más misterioso aún es a partir de qué evolucionaron las plumas. Cabe pensar que se derivan de las mismas células que dieron lugar a las escamas de los reptiles, pero no todo el mundo concuerda en ello.
Pese a todas las incógnitas, podemos conjeturar cómo la selección natural condujo a las aves modernas. Los primeros dinosaurios carnívoros evolucionaron hacia unas patas delanteras y manos más largas, lo que probablemente los ayudara a agarrar y manipular sus presas. Este tipo de agarre habría favorecido la evolución de músculos que permitieran extender con rapidez las patas delanteras y recogerlas, justo el tipo de movimiento que se utiliza para batir las alas en el vuelo activo. Luego vino el recubrimiento del cuerpo con plumas, probablemente como aislante. Dadas estas innovaciones, hay al menos dos vías por las que pudo evolucionar el vuelo. La primera es la conocida como «árboles abajo». Hay indicios para creer que algunos terópodos vivían, al menos en parte, en los árboles. Unos miembros recubiertos de plumas habrían ayudado a estos reptiles a planear de un árbol a otro, o de un árbol al suelo, lo que les hubiera servido para escapar de sus depredadores, encontrar alimento más fácilmente o amortiguar la caída.
Un escenario distinto, y más probable, es la teoría conocida como «suelo arriba»; de acuerdo con ella, el vuelo habría evolucionado a consecuencia de las carreras y saltos con los brazos abiertos que los dinosaurios con plumas debían realizar para capturar a sus presas. Unas alas más largas también podrían haber evolucionado como una ayuda para correr. La perdiz de Chukar, un ave de caza estudiada por Kenneth Dial, de la Universidad de Montana, representa un ejemplo vivo de este paso. Estas perdices no vuelan casi nunca, pero baten sus alas sobre todo como ayuda para correr cuesta arriba. No obtienen así una propulsión adicional, sino más tracción contra el terreno. Los pollos recién nacidos pueden correr por pendientes de 45 grados, y los adultos pueden ascender pendientes de 105 grados, ¡salientes más que verticales!, con sólo correr y batir las alas. La ventaja evidente es que trepar cuesta arriba los ayuda a escapar de sus depredadores. El siguiente paso en la evolución del vuelo sería dar pequeños saltos y mantenerse en el aire brevemente, como hacen los pavos y las codornices para escapar de un peligro.
Tanto en el escenario de «árboles abajo» como el de «suelo arriba», la selección natural podría haber comenzado a favorecer a los individuos que pudieran volar más lejos en lugar de limitarse a planear, saltar o hacer vuelos cortos. Luego vendrían las otras innovaciones compartidas por las aves actuales, como los huesos huecos para ser más ligeras y la quilla.
Aunque podemos especular sobre los detalles, la existencia de fósiles de transición y la evolución de las aves desde los reptiles son hechos. Fósiles como Archaeopteryx y sus parientes posteriores muestran una mezcla de caracteres de aves y reptiles primitivos, y aparecen en el momento esperado del registro fósil. Los científicos predijeron que las aves habían evolucionado a partir de los dinosaurios terópodos y, efectivamente, hemos descubierto dinosaurios terópodos con plumas. Vemos una progresión en el tiempo desde los primeros terópodos con el cuerpo recubierto por estructuras finas y filamentosas a los posteriores con plumas, probablemente buenos planeadores. Lo que vemos en la evolución de las aves es la conversión de unos caracteres antiguos (patas delanteras con dedos y pequeños filamentos en la piel) en otros nuevos (alas sin dedos y plumas), justo como predice la teoría de la evolución.
De vuelta al agua: la evolución de las ballenas
Duane Gish, un creacionista norteamericano, es bien conocido por sus animados y populares (aunque radicalmente desacertados) ataques a la evolución. En una ocasión asistí a una de sus conferencias, durante la cual Gish se burló de la teoría de los biólogos según la cual las ballenas descienden de animales terrestres emparentados con las vacas. ¿Cómo, se preguntaba, podía producirse tal transición si la forma intermedia habría estado mal adaptada tanto en la tierra como en el mar, y por consiguiente no podía explicarse por la selección natural? (Este argumento recuerda al de la media ala de las aves.) Para ilustrar lo que estaba diciendo, Gish proyectó una dispositiva en la que se mostraba una caricatura de sirena con la parte delantera de vaca y la trasera de pez. Aparentemente desconcertada por su propio destino evolutivo, esta bestia claramente inadaptada permanecía en la ribera del agua, con un gran signo de interrogación sobre su cabeza. El dibujo tuvo el efecto buscado: la audiencia rompió a reír. ¿De cuánta estulticia eran capaces los evolucionistas?
Sin duda una «vaca-pez» es un ejemplo grotesco de una forma de transición entre los mamíferos terrestres y los acuáticos, un ejemplo de «evolución en vacarrota»,[12] que diría Gish. Pero dejemos a un lado la retórica y los chascarrillos y observemos la naturaleza. ¿Podemos encontrar algún mamífero que viva tanto en la tierra como en el agua, el tipo de animal que supuestamente no puede haber evolucionado?
Fácil. Un buen candidato es el hipopótamo, que aunque estrechamente emparentado con los mamíferos terrestres, es tan acuático como puede serlo un mamífero terrestre. (Hay dos especies de hipopótamo, el pigmeo y el «normal», cuyo nombre científico es, muy apropiadamente, Hippopotamus amphibius.) Los hipopótamos pasan la mayor parte de su tiempo sumergidos en los ríos y pantanos de los trópicos, explorando sus dominios con ojos, narinas y orejas situados en la parte superior de su cabeza, y todos los cuales pueden cerrarse herméticamente bajo el agua. Los hipopótamos se aparean bajo el agua, y su prole, que nada antes que camina, nada y mama bajo el agua. Al ser sobre todo acuáticos, poseen adaptaciones especiales para salir del agua y pastar: suelen comer por la noche y, como son propensos a quemarse la piel, secretan un fluido rojizo y aceitoso que contiene un pigmento, el ácido hiposudórico, que actúa como un protector solar y posiblemente como antibiótico. Esto ha dado origen al mito de que los hipopótamos sudan sangre. Es evidente que están bien adaptados a su medio, y no es difícil de entender que si encontrasen suficiente alimento bajo el agua, podrían evolucionar hasta convertirse en un animal totalmente acuático como las ballenas.
Pero no tenemos por qué contentarnos con imaginar cómo podrían haber evolucionado las ballenas extrapolando lo que vemos en especies vivas. Disponemos para las ballenas de un excelente registro fósil, gracias a sus hábitos acuáticos y sus huesos robustos que se fosilizan con facilidad. Durante los últimos veinte años hemos podido hacernos una buena idea de cómo evolucionaron. Se trata, además, de uno de nuestros mejores ejemplos de transición evolutiva, ya que disponemos de una serie de fósiles ordenada cronológicamente, quizá un linaje de antecesores a descendientes, que muestra cómo pasaron de la tierra al agua.
Se sabe y acepta desde el siglo XVII que las ballenas y sus primos los delfines y las marsopas son mamíferos. Son animales de sangre caliente, paren sus crías y las amamantan, y tienen pelo alrededor de sus orificios nasales. Los análisis de ADN y la presencia de caracteres vestigiales, como su pelvis y patas traseras rudimentarias, ponen de manifiesto que sus antepasados eran terrestres. Las ballenas casi con seguridad evolucionaron a partir de una especie de artiodáctilo, el grupo de mamíferos con un número par de dedos, como los camellos y los cerdos.[13] Los biólogos creen en la actualidad que el pariente más cercano de las ballenas es, como algún lector ya habrá imaginado, el hipopótamo, así que la evolución imaginada de los hipopótamos a las ballenas no es tan inverosímil, después de todo.
Pero las ballenas tienen sus propias características únicas que las distinguen de sus parientes terrestres. Entre éstas se incluyen la ausencia de patas traseras, las patas delanteras con forma de aleta, la cola aplanada y lobulada, el espiráculo (un orificio nasal en la parte superior de la cabeza), el cuello corto, los dientes simples y cónicos (distintos de los dientes complejos con varias cúspides de los animales terrestres), caracteres especiales de la oreja que les permiten oír bajo el agua, y proyecciones robustas encima de las vértebras para el anclaje de los poderosos músculos natatorios de la cola. Gracias a una fabulosa serie de hallazgos fósiles en Oriente Medio, podemos seguir la evolución de cada uno de estos caracteres (salvo por la cola sin huesos, que no se fosiliza) desde las formas terrestres hasta las formas acuáticas.
Disponemos de abundantes fósiles de mamíferos de hace 60 millones de años, pero ninguno es de ballena. No aparecen hasta unos 30 millones de años después unos animales parecidos a las ballenas actuales. Debería ser posible encontrar formas de transición durante este intervalo. Y, una vez más, es precisamente ahí donde los encontramos. La Figura 12 muestra, en orden cronológico, algunos de los fósiles que participaron en esta transición, que se extiende desde hace 52 millones de años hasta hace 40 millones de años.
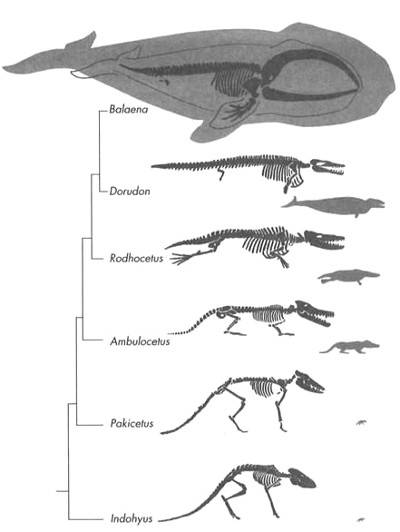
FIGURA 12. Formas de transición en la evolución de las ballenas modernas. (Balaena es la ballena barbada actual, con pelvis y patas traseras vestigiales, mientras que los otros géneros son fósiles de transición.) A la derecha, en gris, se muestra el tamaño relativo de los animales. El «árbol» muestra las relaciones evolutivas entre estas especies.
No hay necesidad de describir esta transición con todo detalle porque los dibujos hablan por sí mismos (si no gritan) de cómo un animal terrestre se echó al agua. La secuencia comienza con un fósil descubierto recientemente de un pariente cercano de las ballenas, un animal del tamaño de un mapache llamado Indohyus. Este animal vivió hace 48 millones de años y, tal como se había predicho, era un artiodáctilo. Es claramente un pariente cercano de las ballenas porque posee rasgos especiales en las orejas y los dientes que sólo se encuentran en las ballenas y en sus antepasados acuáticos. Aunque Indohyus aparece algo más tarde que los antepasados mayormente acuáticos de las ballenas, es probable que se acerque mucho al aspecto que debía de tener el antepasado de los cetáceos. Además, era al menos parcialmente acuático. Lo sabemos porque sus huesos eran más densos que los de los mamíferos completamente terrestres, lo que impedía que flotase en el agua, y porque los isótopos extraídos de sus dientes demuestran que absorbía mucho oxígeno del agua. Lo más probable es que vadeara los ríos o lagos someros para rozar la vegetación o escapar de sus enemigos, de modo parecido a como lo hace en nuestros días un animal parecido, el ciervo-ratón acuático.[14] Esta vida parcialmente acuática probablemente situara a los antecesores de las ballenas en el camino hacia la vida plenamente acuática.
Indohyus no fue un antepasado directo de las ballenas, pero casi con certeza fue primo de uno de ellos. Sin embargo, si nos movemos unos 4 millones de años atrás, hasta hace 52 millones de años, encontramos lo que bien podría ser ese antecesor. Se trata de un cráneo fósil de un animal del tamaño de un lobo llamado Pakicetus, que es un poco más parecido al de las ballenas que el de Indohyus, pues tiene dientes simples y orejas semejantes a las de los cetáceos actuales. Pakicetus todavía no se parecía en nada a una ballena moderna, y en el caso de haberlo podido ver, no se nos ocurriría que él o alguno de sus parientes hubieran de dar origen a una extraordinaria radiación evolutiva. Luego le siguen, en rápida secuencia, una serie de fósiles que con el tiempo fueron haciéndose cada vez más acuáticos. De hace 50 millones de años tenemos el notable Ambulocetus (literalmente, «ballena caminadora»), con un cráneo alargado y reducido pero que todavía tiene miembros robustos que acaban en unas pezuñas que revelan su ascendencia. Probablemente pasara casi todo el tiempo en aguas poco profundas, y sobre la tierra firme se habría bambaleado con dificultad, como les pasa a las focas. Rodhocetus (hace 47 millones de años) es todavía más acuático. Sus narinas han migrado un poco hacia atrás, y su cráneo es más alargado. A juzgar por sus robustas proyecciones en la columna vertebral para anclar los músculos de la cola, Rodhocetus debió de ser un buen nadador, mientras que el pequeño tamaño de su pelvis y extremidades posteriores debían suponerle una desventaja en tierra firme. No cabe duda que debía pasar casi todo el tiempo en el mar. Por último, hace 40 millones de años encontramos los fósiles Basilosaurus y Dorudon, que son claramente mamíferos acuáticos con el cuello corto y los orificios nasales en la parte superior del cráneo. Sabemos que no podían pasar ningún tiempo en tierra firme por su pelvis y extremidades posteriores de tamaño reducido (apenas medio metro en Dorudon, un animal de 15 metros) y desvinculadas del resto del esqueleto.
La evolución de las ballenas a partir de animales terrestres se produjo con notable rapidez: los desarrollos más importantes se produjeron en apenas 10 millones de años. Eso no es mucho más de lo que nos costó a nosotros divergir de nuestro antepasado común con los chimpancés, una transición que involucró muchas menos modificaciones en el cuerpo. No obstante, debe tenerse en cuenta que la adaptación a la vida en el mar no comportó la evolución de ningún carácter nuevo, sólo modificaciones de caracteres antiguos.
Pero ¿por qué algunos animales volvieron al agua? Después de todo, unos cuantos millones de años atrás habían invadido los continentes. No estamos seguros de por qué se produjo una migración inversa, pero tenemos varías ideas. Una posibilidad contempla la desaparición de los dinosaurios junto a sus feroces parientes marinos, los monosauros, ictiosauros y plesiosauros, todos ellos depredadores de peces. Estos animales no sólo hubieran competido por el alimento con los mamíferos acuáticos, sino que probablemente los hubieran convertido en sus presas. Una vez extinguidos sus competidores reptilianos, los antepasados de las ballenas debieron de encontrar un nicho disponible, libre de depredadores y repleto de alimento. El mar se ofrecía para la invasión, y todos sus beneficios estaban a pocas mutaciones de distancia.
Qué dicen los fósiles
Si llegados a este punto el lector se siente abrumado por los fósiles, le consolará saber que he omitido otros centenares que también muestran la evolución. Está la transición entre los reptiles y los mamíferos, tan ampliamente documentada con «reptiles mamiferoides» que constituye el asunto de varios libros. Están también los caballos, una ramificación evolutiva que parte de unos pequeños antepasados de cinco dedos a la orgullosa especie de pezuña que vemos hoy. Y, naturalmente, el registro fósil de los humanos, descrito en el capítulo 8, que sin lugar a dudas constituye el mejor ejemplo de una predicción evolutiva que los descubrimientos han satisfecho.
A riesgo de aburrir al lector, mencionaré brevemente unas pocas formas de transición más. La primera corresponde a un insecto. Sobre la base de sus semejanzas anatómicas, los entomólogos habían postulado que las hormigas debían de haber evolucionado a partir de unas avispas no sociales. En 1967, E. O. Wilson y colaboradores hallaron, preservada en ámbar, una hormiga «transicional» que poseía casi el catálogo completo de caracteres de hormiga y avispa que los entomólogos habían predicho (Figura 13).
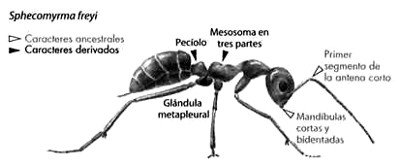
FIGURA 13. Insecto transicional, una de las primeras hormigas, con caracteres primitivos de avispa (el grupo ancestral predicho) y caracteres derivados propios de las hormigas. De esta especie, Sphecomyrma freyi, se encontró un único espécimen preservado en ámbar de hace 92 millones de años.
De modo parecido, desde hacía mucho tiempo se suponía que las serpientes habían evolucionado a partir de reptiles con aspecto de lagarto que perdieron las patas, ya que los reptiles con patas aparecen en el registro fósil mucho antes que las serpientes. En 2006, unos paleontólogos que excavaban en Patagonia encontraron un fósil de la serpiente más antigua conocida, de hace 90 millones de años. Tal como se había predicho, tenía la cintura pélvica pequeña y las patas posteriores reducidas. Pero quizá el hallazgo más emocionante sea un fósil de hace 530 millones de años descubierto en China llamado Haikouella lanceolata, que se asemeja a una pequeña anguila con una aleta dorsal con volantes. Pero también tenía cabeza, cerebro, corazón y una cuerda cartilaginosa que le recorría el dorso: un notocordio. Este fósil marca lo que quizá sea el primer cordado, el grupo que dio origen a todos los vertebrados, incluidos los humanos. En este complejo organismo de apenas un par de centímetros de longitud podrían encontrarse las raíces de nuestra propia evolución.
El registro fósil nos enseña tres cosas. En primer lugar, nos habla de la evolución con una voz poderosa y elocuente. El registro de las rocas confirma varias de las predicciones de la teoría de la evolución: el cambio gradual dentro de los linajes, la división de linajes y la existencia de formas de transición entre tipos de organismos muy distintos. No es posible ignorar toda esta evidencia, no es posible arrumbarla. La evolución ocurrió, y en muchos casos sabemos cómo.
En segundo lugar, cuando hallamos formas de transición, las encontramos dentro del registro fósil justo allí donde las esperamos. Las primeras aves aparecen después de los dinosaurios pero antes que las aves modernas. Hallamos ballenas ancestrales que llenan la brecha entre sus propios antepasados, pobres marineros, y las ballenas claramente modernas. Si la evolución no fuese cierta, no encontraríamos los fósiles en un orden con sentido evolutivo. Interrogado sobre la observación que podría refutar la evolución, el biólogo cascarrabias J. B. Haldane al parecer gruñó: «¡Fósiles de conejo en el Precámbrico!». (El período geológico que acabó hace 543 millones de años.) Huelga decir que jamás se han hallado conejos precámbricos ni ningún otro fósil anacrónico.
Por último, el cambio evolutivo, incluso el más radical, casi siempre implica una remodelación de lo viejo en lo nuevo. Las patas de los animales terrestres son variaciones de los robustos miembros de los peces ancestrales. Los minúsculos huesecillos del oído medio de los mamíferos están hechos a partir de huesos mandibulares de sus antecesores reptilianos. Las alas de las aves derivan de las patas de los dinosaurios. Y las ballenas son animales terrestres alargados cuyas extremidades anteriores se han convertido en aletas y cuyas narinas han migrado hasta la parte superior de la cabeza.
No hay razón alguna para que un diseñador celeste que crea unos organismos desde la nada, igual que un arquitecto diseña sus edificios, hiciera las nuevas especies remodelando caracteres de otras especies existentes. Podía haber construido cada especie desde cero. Pero la selección natural sólo puede actuar modificando lo que ya existe, no puede producir caracteres nuevos a partir de la nada. El darwinismo predice, por consiguiente, que las especies nuevas deben ser modificaciones de las existentes. El registro fósil confirma abundantemente esta predicción.